
No todos los animales cuentan con lo que puede considerarse, con propiedad, un sistema circulatorio. Y muchos animales, aunque tienen bomba o bombas de impulsión, tienen sistemas circulatorios abiertos. En los sistemas abiertos la sangre o hemolinfa bombeada se vierte a una amplia cavidad interna a través de un sistema de arterias que se ramifican de forma sucesiva. Luego es recogida por un sistema de venas equivalente al anterior y devuelto a la bomba de impulsión. Los sistemas abiertos ejercen un menor control que los cerrados sobre el flujo de sangre a los diferentes órganos, aunque en cualquier caso las demandas de los tejidos son satisfechas de manera conveniente.
El sistema abierto más complejo que se conoce es el de algunos crustáceos, que alcanzan un desarrollo muy similar al de los sistemas cerrados. El sistema arterial llega hasta el final de las extremidades, la hemolinfa se vierte en pequeños espacios abiertos y de ellos es recogida por unos senos a modo de pequeños canales que la devuelven al corazón, no sin antes pasar por las branquias. Así pues, en estos crustáceos la sangre se oxigena en su trayecto de vuelta a la bomba de impulsión.
Las cosas son diferentes en los animales que tienen sistemas cerrados. Los cefalópodos son los únicos moluscos con sistema cerrado. En estos la sangre también pasa por las branquias antes de alcanzar el corazón sistémico o corazón principal, de donde se distribuye al resto de órganos y tejidos. A diferencia de los crustáceos, los moluscos cefalópodos tienen dos corazones branquiales que bombean la sangre venosa procedente de los tejidos a las branquias. Así se oxigena y se descarga de CO2 antes de llegar al corazón sistémico. Una característica notable de este sistema es que al trabajar con sangre venosa, los corazones auxiliares trabajan en condiciones virtualmente anóxicas, con muy escaso aporte de oxígeno.
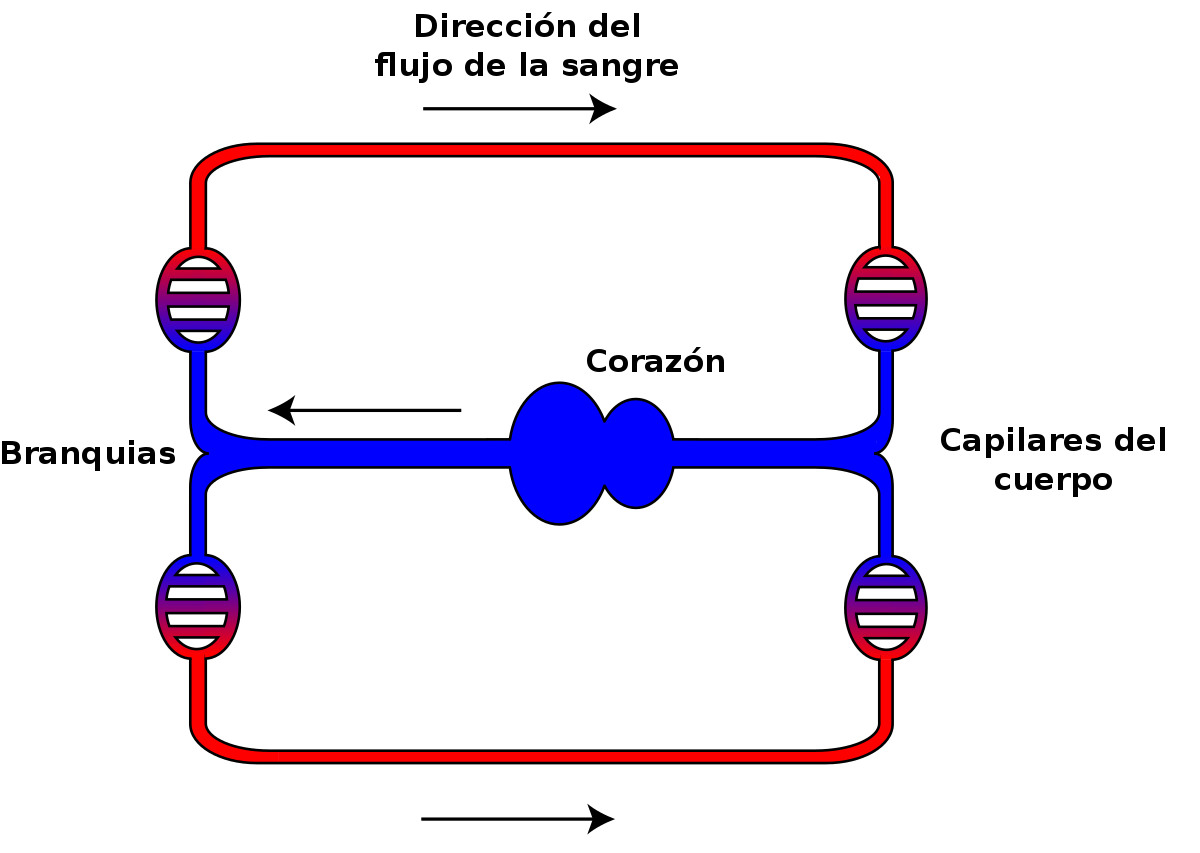
Los sistemas circulatorios cerrados más desarrollados son los de los vertebrados, y hay una gran variedad de disposiciones anatómicas en este grupo. Lo más probable es que los de los primeros vertebrados consistieran en un circuito muy simple que siguiera, más o menos, la secuencia: corazón, arteria, branquias, aorta, arterias, arteriolas, capilares, vénulas, venas y corazón. Este sistema es diferente de los anteriores porque en aquellos la sangre circulaba de las branquias al corazón y en este circula del corazón a las branquias. Es un buen dispositivo a efectos de oxigenar la sangre, pero reduce mucho la velocidad de circulación desde el corazón a los tejidos, porque al pasar por los capilares branquiales, estos, por su pequeño calibre, ofrecen una gran resistencia al paso de la sangre y la ralentizan. Los peces mantienen las características generales de este sistema, aunque en algunos casos incorporan alguna bomba auxiliar antes del corazón sistémico.
La transición al medio terrestre trajo importantes novedades. La aparición de órganos respiratorios distintos de las branquias complicó de forma notable la arquitectura del circuito. Como vimos aquí, hay peces que son capaces de respirar en aire a través de estructuras tales como la cavidad bucal, el intestino, la vejiga natatoria o la cloaca. En los primeros que desarrollaron esa capacidad el órgano respiratorio recibe sangre del circuito sistémico, lo que da lugar a que, una vez oxigenada, se mezcle con la desoxigenada que procede del resto de los tejidos antes de entrar en el corazón. Por lo tanto, cuando respiran en aire los tejidos sistémicos reciben sangre parcialmente oxigenada, porque las branquias, en caso de ser funcionales, aportan muy poco oxígeno. Esa particularidad, que ha sido tradicionalmente considerada una desventaja de esos peces, tiene sin embargo una ventaja por comparación con los que respiran en agua a través de las branquias: como la sangre que llega al corazón está parcialmente oxigenada, el corazón toma de ella el oxígeno que necesita.
A diferencia de los anteriores, en los peces pulmonados –y en concreto, en el respirador aéreo obligado Protopterus aethiopicus– la sangre procedente de los pulmones no se mezcla con la procedente de los tejidos antes de entrar en el atrio (aurícula) del corazón, sino que entra en este directamente. Y por otro lado, la configuración anatómica del corazón de estos peces es tal que permite que la sangre procedente de los pulmones y la que llega del resto de los tejidos solo se mezclen en una pequeña proporción, de manera que la que viene de los pulmones se dirige preferentemente a la circulación sistémica, y la que procede de los tejidos a los pulmones. Aunque anatómicamente no hay una separación clara entre los dos circuitos, el sistémico o general, y el pulmonar, funcionalmente se produce una cierta diferenciación, lo que constituye un anticipo de los sistemas totalmente diferenciados que aparecieron posteriormente en otros vertebrados.
Los anfibios que respiran en aire tienen ya dos cámaras auriculares separadas. La sangre oxigenada procedente de los pulmones entra en la aurícula izquierda, mientras la sangre sistémica venosa entra en la derecha a través del seno venoso. Ambas sangres están separadas hasta que llegan al ventrículo. Pero, gracias a las características ventriculares, tampoco en este se mezclan en una medida significativa. En anfibios, además, hay que introducir la irrigación de la piel en el análisis, dado que puede contribuir de forma significativa al intercambio de gases. El caso de la rana toro (Rana catesbeiana) es muy ilustrativo. Prácticamente toda la sangre oxigenada procedente de los pulmones llega a la aurícula izquierda y sale hacia la circulación sistémica, se descarga de O2 en los tejidos y vuelve al corazón, donde entra por la aurícula derecha; la mayor parte de esa sangre venosa desoxigenada se dirige a la vía pulmo-cutánea (dos tercios a los pulmones, y el otro tercio a la piel). Lo curioso es que la sangre (oxigenada) que procede de la piel se mezcla con la procedente de los tejidos antes de llegar al corazón.

Los reptiles no cocodrilianos (tortugas, serpientes y lagartos) tienen el ventrículo parcialmente separado en tres cámaras, y aunque la separación no es total, también en estos hay poca mezcla de sangre oxigenada y desoxigenada. No se conoce bien el mecanismo, pero las arterias pulmonares reciben preferentemente sangre desoxigenada. En los cocodrilianos el ventrículo está completamente dividido en dos cámaras por un septo. Del ventrículo izquierdo sale la arteria pulmonar y una arteria sistémica, la izquierda, y del ventrículo izquierdo sale la aorta sistémica derecha; sendas válvulas controlan el flujo a través de ambas aortas y están, además, conectadas nada más salir del corazón. Ese sofisticado dispositivo no solo propicia una distribución selectiva de la sangre desoxigenada a los pulmones prácticamente perfecta, sino que durante el buceo permite limitar el flujo de sangre a los pulmones cuando estos se han quedado sin oxígeno.
Los diferentes dispositivos vistos en peces (de respiración aérea), anfibios y reptiles son funcionales en animales que no tienen garantizado un alto contenido en oxígeno en sus pulmones de manera permanente. Permiten dirigir la sangre desoxigenada y oxigenada hacia diferentes destinos dependiendo de las circunstancias; por ejemplo, dependiendo de la disponibilidad ambiental de O2 y, por ello, de su concentración en el órgano respiratorio. Por eso, la adquisición de un doble sistema, pulmonar y sistémico, por parte de aves y mamíferos no debe entenderse como un “último” paso en la evolución de los sistemas respiratorios hacia una hipotética configuración ideal. La disposición en serie del circuito sistémico y del pulmonar obliga a que los flujos hacia uno y el otro sean idénticos. Ello impide generar flujos diferentes en un y otro circuito, pero en aves y mamíferos no hay tal necesidad, pues, como se ha dicho, estos grupos tienen elevadas concentraciones de oxígeno en sus pulmones de forma permanente.
Por otro lado, esa configuración permite generar presiones hidrostáticas muy diferentes en cada circuito: en el sistémico son necesarias altas presiones para vencer la resistencia que ofrece el sistema capilar, pero en el pulmonar la presión no ha de ser muy alta, pues de lo contrario saldría plasma sanguíneo hacia los espacios extracelulares e, incluso, al interior de los alveolos, lo que aumentaría peligrosamente la longitud de la vía de difusión. Como consecuencia de esa disposición en serie, ambos circuitos se mantienen completamente separados en el corazón. En animales con muy altas demandas metabólicas, como aves y mamíferos, eso es ventajoso pues permite garantizar que sangres con diferentes contenidos de O2 no se mezclan en ningún momento, lo que favorece el transporte de oxígeno a los tejidos. Pero que ese dispositivo resulte muy ventajoso para aves y mamíferos no quiere decir que los animales que carecen de él tengan, a esos efectos, una configuración incompleta. Como hemos visto, el resto de vertebrados que respiran en aire en la práctica también han conseguido evitar la mezcla de sangres, aunque no de forma completa, y disponen, además de otras funcionalidades de las que carecen los dos grupos homeotermos.
Nota:
En las siguientes anotaciones hemos tratado otros aspectos de los sistemas circulatorios que complementan lo expuesto aquí:
La distribución del agua animal y el curioso caso del potasio
Un viaje a través del sistema circulatorio humano
El flujo sanguíneo se reorganiza en respuesta a las necesidades
Sobre el autor: Juan Ignacio Pérez (@Uhandrea) es catedrático de Fisiología y coordinador de la Cátedra de Cultura Científica de la UPV/EHU

Paco
Hecho de menos más gráficos explicativos. Lo suyo sería uno por cada sistema concreto que se habla.
Buen artículo en todo caso.
Corazones miogénicos y neurogénicos – Cuaderno de Cultura Científica
[…] 2Aquí puede consultarse una descripción muy somera de los diferentes configuraciones cardiacas en vertebrados. […]