Atraer polinizadores: el calor precedió al color de las flores
La mayor parte de las angiospermas (plantas con flores) depende de los insectos y de otros animales para su polinización. Por este motivo, las angiospermas han desplegado variadísimas formas para atraer a estos polinizadores y lograr que transporten el polen de unas flores a otras. Estas estrategias incluyen colores vivos, comprendiendo regiones del espectro electromagnético no visible para nosotros (ultravioleta), olores y recompensas en forma de néctar.
Las angiospermas comenzaron su diversificación en el Cretácico temprano, hace unos 130-140 millones de años. Sin embargo, la polinización por insectos ya se producía mucho antes en plantas gimnospermas, carentes de flores. En 2015, científicos españoles describieron moscas conservadas en ámbar que mostraban un aparato bucal adaptado a la absorción de néctar y que estaban asociadas a polen de gimnospermas, probablemente del orden Bennettitales. Otras evidencias indirectas sugieren que ya existían insectos polinizadores en el Jurásico superior, hace 163 millones de años. ¿Cómo eran atraídos los insectos hacia plantas carentes de flores?
La hipótesis que se maneja desde hace años se basa en la termogénesis, producción de calor en las estructuras reproductivas de algunas plantas. Esta termogénesis se produce por el mismo mecanismo de producción de calor de los animales endotermos (aves y mamíferos). Este fenómeno se produce en las mitocondrias cuando la energía química de los nutrientes no se almacena en el ATP, y se disipa en forma de calor. La termogénesis vegetal se ha observado en los conos o estróbilos de dos familias de gimnospermas (cicadáceas y zamiáceas), muy primitivas desde el punto de vista evolutivo. El origen de las cicadáceas, cuyo representante más conocido es la falsa palmera (género Cycas), se remonta al Pérmico, hace 300 millones de años.
También se produce calor en las flores de algunas familias de angiospermas, como varias especies de aráceas, las magnolias o las chirimoyas. En este vídeo podemos ver como el calor desprendido por la flor de la arácea Symplocarpus foetidus le permite derretir la nieve y salir al exterior. El calor también ayuda a difundir los insoportables olores a carne podrida de otra arácea, la extraordinaria “flor cadáver” (Amorphophallus titanium). Aquí pueden encontrar una excelente revisión del tema, realizada por investigadores españoles.
El calor desprendido en todos estos casos produce aumentos de temperatura diversos, que van desde 1ºC a 4ºC sobre la temperatura ambiente en chirimoyas y magnolias, hasta 20ºC en las cicadáceas y 35ºC en algunas especies de Philodendron. Se pensaba que este calor, además de ofrecer un ambiente agradable a los insectos, particularmente a los coleópteros, promueve la volatilización de sustancias atractivas para los polinizadores.
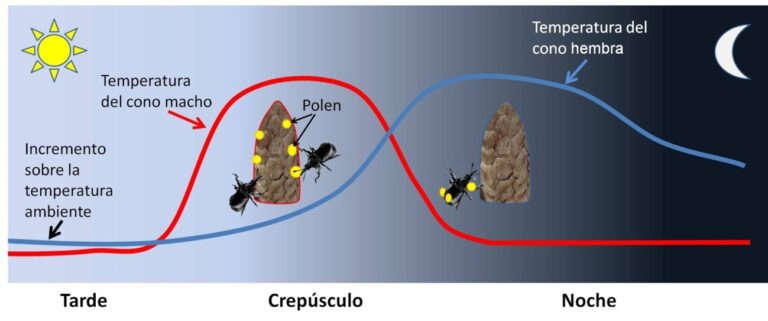
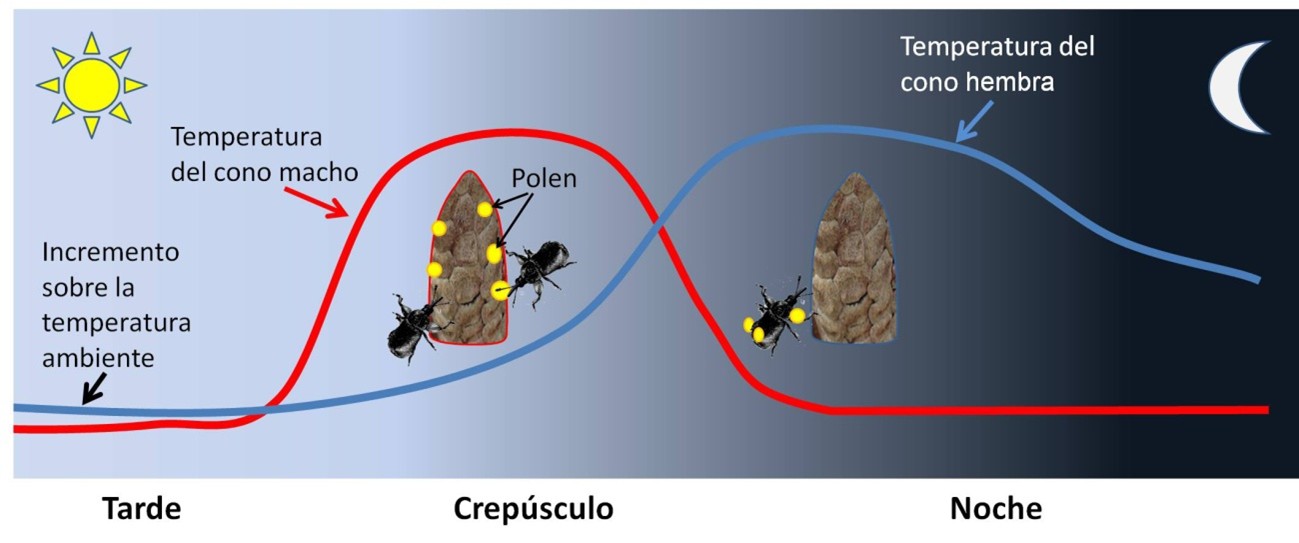
Un estudio que acaba de ser publicado en la revista Science por un grupo internacional, coordinado desde la universidad de Harvard, nos permite conocer con mucho más detalle lo que pudo ser el proceso ancestral de polinización por parte de los insectos. La investigación se centró en una planta gimnosperma, la Zamia furfuracea y su polinizador habitual, el pequeño escarabajo Rhopalotria furfuracea (Figura 1). Los conos de Zamia suelen incrementar su temperatura al atardecer, manteniéndose entre 3ºC y 6ºC por encima de la temperatura del aire.

Los escarabajos se marcaron con un producto fluorescente que permitió seguir sus movimientos en la oscuridad. Sorprendentemente, los conos masculinos, productores de polen, aumentaban la temperatura antes que los femeninos, atrayendo a los escarabajos. Luego se enfriaban, al tiempo que los femeninos incrementaban su producción de calor. Esto hacía que los escarabajos, que se habían cargado de polen, se dirigieran a los conos hembra y fecundaran a los gametos femeninos (Figura 2).
Los investigadores comprobaron mediante varios experimentos que los escarabajos eran sensibles a la radiación infrarroja que emiten los cuerpos calientes. De hecho, eran atraídos por conos artificiales con temperatura similar a la de los naturales. Esta sensibilidad se localizaba en el extremo de las antenas. Cuando esta zona fue microdiseccionada los escarabajos dejaron de dirigirse a los conos calientes.
Resultó muy curioso comprobar que en las células sensoriales del extremo de las antenas se expresaba el receptor TRPA1, un canal iónico de la membrana celular que también está implicado en la termorrecepción de mosquitos y serpientes.
Lo más intrigante del estudio es el desfase en el incremento de temperatura entre los conos masculinos y femeninos. Se ha podido comprobar que este desfase responde a un reloj circadiano similar al que regula otros procesos con periodicidad diaria en plantas y animales, pero lo sorprendente es que conos masculinos y femeninos manejen relojes independientes.
Todos estos nuevos resultados hacen suponer que la polinización de plantas anteriores a las angiospermas utilizó como señal el calor y la detección de la consecuente radiación infrarroja por parte de coleópteros nocturnos. Cuando se diversificaron las plantas con flores, la gran diversidad de estímulos cromáticos expandió la nómina de polinizadores a los insectos voladores y diurnos, sobre todo himenópteros y lepidópteros. La nueva y creciente red de relaciones insectos/plantas pudo impulsar la diversificación evolutiva de las angiospermas hasta las más de 350 000 especies conocidas actualmente. En cambio, gimnospermas como las cicadáceas o zamiáceas (375 y 111 especies, respectivamente) pudieron quedar restringidas desde el punto de vista evolutivo por la menor diversidad de sus sistemas de atracción de polinizadores.
Referencias
Valencia-Montoya, W.A., Liénard, M.A., Rosser, N. et al. (2025). Infrared radiation is an ancient pollination signal. Science 390(6778):1164-1170. doi: 10.1126/science.adz1728.
Sobre el autor: Ramón Muñoz-Chápuli Oriol es Catedrático de Biología Animal (jubilado) de la Universidad de Málaga.