Sistemas nerviosos: evolución de la estructura encefálica

J. B. Johnston, uno de los fundadores de la neurobiología evolutiva, afirmó en 1923, refiriéndose a diferentes grupos de vertebrados, que “new structures have not appeared” (no han aparecido nuevas estructuras). Y sin embargo, tal y como hemos visto en las anotaciones anteriores, los encéfalos de unas especies carecen de estructuras que sí aparecen en otras. Es más, el uso del prefijo “neo” en neuroanatomía es muy común.
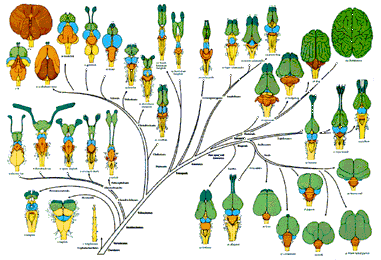
Ludwig Edinger, otro de los precursores, había propuesto en 1908 que en el curso de la evolución del encéfalo de vertebrados se añadió un “neencephalon” al “palaeencephalon” preexistente. Esa idea tuvo amplia aceptación en la primera mitad del siglo XX y dio lugar a la visión más general de que los encéfalos han evolucionado por la acumulación de partes nuevas. La idea tiene el atractivo de que implica un aumento de la complejidad: como se añaden partes nuevas, el número de regiones aumenta, luego la complejidad es mayor. Sin embargo, Johnston había comprobado por sí mismo y a partir de trabajos de otros investigadores que las consideradas nuevas tenían estructuras homólogas1 en los vertebrados “inferiores”. Por ejemplo, el pallium de peces y reptiles, aunque de aspecto muy diferente, es homólogo al neocortex de mamíferos y por lo tanto, éste no se puede considerar propiamente “nuevo”. En la actualidad se acepta que en el curso de la evolución de vertebrados no han aparecido estructuras nuevas, entendiendo nuevas como no homólogas a estructuras precursoras, pero lo que es claro es que, en general, conforme han ido surgiendo nuevos grupos, sus encéfalos han ido siendo cada vez más complejos, porque el número de regiones encefálicas o agrupamientos celulares ha aumentado. De hecho, una somera comparación de su número sugiere que la complejidad ha aumentado y disminuido en la evolución de los vertebrados en varias ocasiones.
Número de áreas encefálicas diferenciables (la secuencia sigue un orden de diferenciación en el curso de la evolución): mixinos (peces brujas): 36; lampreas: 25; tiburones: 46; rayas: 45; bichires de Cuvier (osteíctio actinopterigio): 46; teleósteos: 71; peces pulmonados: 29; salamandras: 31; ranas: 50; reptiles: 85; aves: 87; mamíferos: 257.
A la vista de esos datos parece que la complejidad encefálica aumentó en los peces teleósteos, más adelante en el origen de los amniotas y después con los mamíferos. Y descendió con los peces pulmonados y las salamandras. Así pues, los encéfalos han variado en su número de estructuras y, en general, la tendencia ha sido de aumento.
Hay varios mecanismos que pueden explicar el aumento de regiones encefálicas sin que se hayan generado (filogenéticamente) nuevas estructuras. Pueden haber aparecido por conversión filogenética (cuando una estructura se transforma en otra) o proliferación filogenética, y dentro de esta, segregación (cuando una estructura da lugar a dos o más) y adición (cuando una estructura surge ex novo).
La aparición de estructuras en láminas a partir de una anterior homóloga pero no laminar es un caso paradigmático de conversión filogenética. Tienen estructuras laminares el lóbulo vagal de los teleósteos ciprínidos, el torus semicircularis de los peces eléctricos Gymnotidae, y el núcleo geniculado lateral dorsal de los mamíferos; todas estas estructuras son homólogas a otras no laminadas que existen en grupos relacionados. Una clara ventaja de la estructura laminar es que minimiza las longitudes de conexión neuronal y, por lo tanto, permite ahorros en energía, espacio y tiempos de conducción. También facilita la emergencia de funciones más complejas que los reflejos. La estructura laminar se desarrolla con facilidad y es funcionalmente ventajosa porque las neuronas en láminas diferentes pueden conectarse con mínimo cableado (dendritas y axones).
El término segregación filogenética se acuñó para denominar la diferenciación de ganglios (núcleos) a partir de un tejido poco diferenciado del tallo encefálico, y se puede aplicar, en general, a aquellos casos en los que regiones encefálicas homogéneas dan lugar a varias regiones diferentes, ninguna de las cuales es homóloga (una a una) a la antigua. Un ejemplo es el tálamo dorsal, que contiene bastantes más núcleos en amniotas que en anamniotas. Otro es el del complejo preglomerular de los peces (osteíctios) actinopterigios, con un número de núcleos muy diferente en unas especies y otras. El número de núcleos es mayor cuanto más recientemente han evolucionado las especies. El aumento en el número de núcleos se atribuye a un proceso de proliferación filogenética, que muy probablemente ha ocurrido por segregación.
Los casos mejor conocidos de adición son el lóbulo de la línea lateral electrosensorial de peces eléctricos, la válvula cerebelar, que solo aparece en teleósteos, la porción electrosensorial del torus semicircularis del encéfalo medio de los Gymnotidae y los siluros; o el torus longitudinalis del techo óptico de todos los (osteíctios) actinopterigios. Las adiciones pueden producirse mediante desarrollo de nuevas estructuras a partir de tejido embrionario. Ese tejido precursor, que en estadios evolutivos anteriores daba lugar a una estructura determinada en el encéfalo adulto, puede –además de conservar la anterior- generar nuevas estructuras en fases evolutivas posteriores. Lo puede hacer, además, mediante dos posibles vías. Una consistiría en una nueva forma de expresión génica en una zona de ese tejido precursor. Y la otra se puede producir si el tejido en cuestión alcanza un tamaño mayor y parte de la estructura se ve expuesta a estímulos ontogenéticos diferentes, de manera que su desarrollo cursa de forma distinta, dando lugar a una estructura nueva.
Un aspecto de especial interés es el modo en que se han desarrollado las diferentes áreas corticales. Se ha constatado que mamíferos de muy diferentes grupos tienen las mismas áreas sensoriales primarias y secundarias bien diferenciadas. Y algo similar cabe decir de las áreas motoras en placentarios. Lo más probable es, por lo tanto, que las áreas sensoriales y motoras de niveles básicos de la corteza surgieran ya en los primeros mamíferos, lo que no parece ser el caso de las áreas corticales de niveles superiores. ¿Cómo surgieron, entonces, esas áreas de orden superior? Al respecto se han formulado diferentes hipótesis, como la de la posibilidad de una agregación selectiva de heterogeneidades modulares preexistentes en las áreas antigua, o la duplicación de áreas preexistentes y posterior diferenciación funcional, entre otras. No obstante, hay aspectos sustanciales de la arquitectura encefálica y de la diversificación de áreas que no son explicadas de forma satisfactoria por ninguna de las hipótesis formuladas.
Para concluir, remarcaremos la idea de que sean cuales sean los mecanismos implicados en la aparición de nuevas áreas o regiones en el encéfalo de los vertebrados, parece claro que un mayor número de áreas es ventajoso desde el punto de vista funcional, porque de esa forma pueden especializarse en tareas diferentes. La división del trabajo redunda en una mayor eficiencia aunque también reduce la flexibilidad del sistema. No obstante, en el curso de la evolución de las especies de vertebrados parece haberse favorecido la eficiencia en perjuicio de la flexibilidad. De esa forma se han alcanzado encéfalos de gran tamaño, lo que no hubiera sido posible con bajos niveles de diferenciación. En síntesis, a lo largo de la evolución se han ido desarrollando encéfalos más complejos porque son funcionalmente útiles y han sido la única forma de tener grandes números de neuronas sin sacrificar su funcionalidad.
Fuentes:
Eric R, Kandel, James H. Schwartz, Thomas M. Jessell, Steven a. Siegelbaum & A. J. Hudspeth (2013): Principles of Neural Science 5th ed., Mc Graw Hill, New York
Georg F. Striedter (2005): Principles of Brain Evolution, Sinauer & Co, Sunderland USA
Nota:
1Homólogo es un rasgo que tiene origen común, o sea que evolucionó una vez.
Sobre el autor: Juan Ignacio Pérez (@Uhandrea) es catedrático de Fisiología y coordinador de la Cátedra de Cultura Científica de la UPV/EHU
Txema M.
Lo primero es felicitarte por la serie neurológica; es un ejemplo de divulgación científica de calidad. Enhorabuena y gracias por tu esfuerzo.
Te pregunto por las dudas que me surgen leyendo uno de los párrafos. Dices que «A la vista de esos datos parece que la complejidad encefálica aumentó en los peces teleósteos, más adelante en el origen de los amniotas y después con los mamíferos. Y descendió con los peces pulmonados y las salamandras. Así pues, los encéfalos han variado en su número de estructuras y, en general, la tendencia ha sido de aumento.» En la tabla que anexas sobre el número de áreas encefálicas diferenciables los teleósteos aparecen con 71 áreas, mientras que los peces pulmonados poseen sólo 29. Me pregunto si el número de 71 áreas es común a todo el grupo de teleósteos o si es tan solo una media. En ese último caso ¿cabría la posibilidad de que los antecesores de los peces pulmonados hubiesen tenido un número reducido de áreas diferenciables, un número menor aún que el de sus descendientes?
¿Respecto a qué antepasados se dió la reducción en el caso de las salamandras? Lo digo porque creía que algún tipo de pez pulmonado se encontraba en el camino evolutivo que llevó a los tetrápodos. Si fuera así, igual no hubo reducción de áreas en las salamandras, al menos en comparación con los peces pulmonados.
Y por último: si se da verdaderamente en el curso de la evolución el caso de los torna atrás que citas en peces pulmonados y salamandras ¿cómo ocurre? ¿Se da una desaparición de estructuras preexistentes o cabe pensar en un proceso de fusión de áreas encefálicas?
Hitos en la red #183 | Enlace Recomendado | Naukas
[…] escribiendo Juan Ignacio Pérez sobre sistemas nerviosos, y esta es su última entrega Sistemas nerviosos: evolución de la estructura encefálica, y este cronista está escribiendo sobre conceptos termodinámicos (clásicos y […]
Juan Ignacio Pérez Iglesias
Hola Txema
Antes de responder, te advierto: no es mi campo; lo que he escrito lo he sacado de lecturas. Y en concreto, las dudas que me planteas se refieren a asuntos tratados en Striedter (2005).
1. El autor no especifica si es un dato medio o un valor común. Habla de recuentos hechos en especies representativas. Me temo que no son datos de gran rigor estadístico, pero sospecho que los datos disponibles para diferentes especies se parecen mucho en un mismo grupo. No creo que los antecesores de los pulmonados tuviesen menos que estos. El autor lo habría especificado. Sospecho que la pérdida de complejidad en estos peces tiene que ver con un modo de vida diferente.
2, No creo que tenga un término de comparación concreto para las salamandras, porque obviamente no hay datos relativos a peces ya extinguidos como los osteolepiformes. Supongo que la comparación la hace con relación al conjunto de datos de peces que figuran en la relación. Si omitimos a las lampreas, que no son ni peces, y a los pulmonados, el dato de las salamandras es inferior a todos los demás, por lo que imagino que el autor infiere que la transición al medio terrestre por esa vía ha conllevado pérdida de complejidad. Hasta donde yo sé (muy poco) los peces pulmonados actuales tienen poco que ver con los anfibios. Los pulmonados son Dipnomorpha y los anfibios provienen de los osteolepiformes y, por lo tanto, de los Tetrapodomorpha, y esas dos vías, dipnomorfos y tetrapodomorfos son independientes y ambas proceden de Rhipidistia.
3. No lo sé.
Lamento no ser de más ayuda.
Y gracias por una lectura tan concienzuda.
Los cerebros de aves y mamíferos evolucionaron diferente
[…] de forma independiente, a pesar de compartir un ancestro común. Estos hallazgos desafían la visión tradicional de la evolución del cerebro y demuestran que, aunque existen funciones cerebrales comparables […]