Relaciones hídricas y salinas de los animales

Los líquidos corporales de los animales se distribuyen en tres espacios o compartimentos, principalmente: celular (líquido intracelular), espacios entre las células (líquido intersticial) y, de haberlas, cavidad general del cuerpo y sistema vascular (líquido celómico, plasma sanguíneo, hemolinfa o equivalente). Como vimos aquí, el líquido intersticial y el plasma pueden ser considerados conjuntamente líquido extracelular (o medio interno). En los animales sin sistema vascular la razón de esa identidad es obvia. En los animales con sistema circulatorio abierto no hay discontinuidad alguna entre la hemolinfa y el líquido que baña las células. Y en los de sistema cerrado, los capilares tienen poros (entre las células endoteliales o, incluso, atravesando tales células) a través de los cuales se establece una continuidad casi absoluta entre ambos sistemas; las únicas sustancias que no atraviesan la barrera que representa el endotelio capilar son ciertas moléculas, como proteínas plasmáticas, cuyo gran tamaño impide su salida del capilar hacia los espacios intercelulares. Por lo tanto, el líquido intersticial y el plasma (o equivalente) tienen la misma concentración osmótica y casi idéntica composición iónica. Sin embargo, aunque los líquidos extra e intracelular tengan también la misma concentración osmótica, la composición de iones y de ciertas moléculas orgánicas es muy diferente en ambos fluidos.
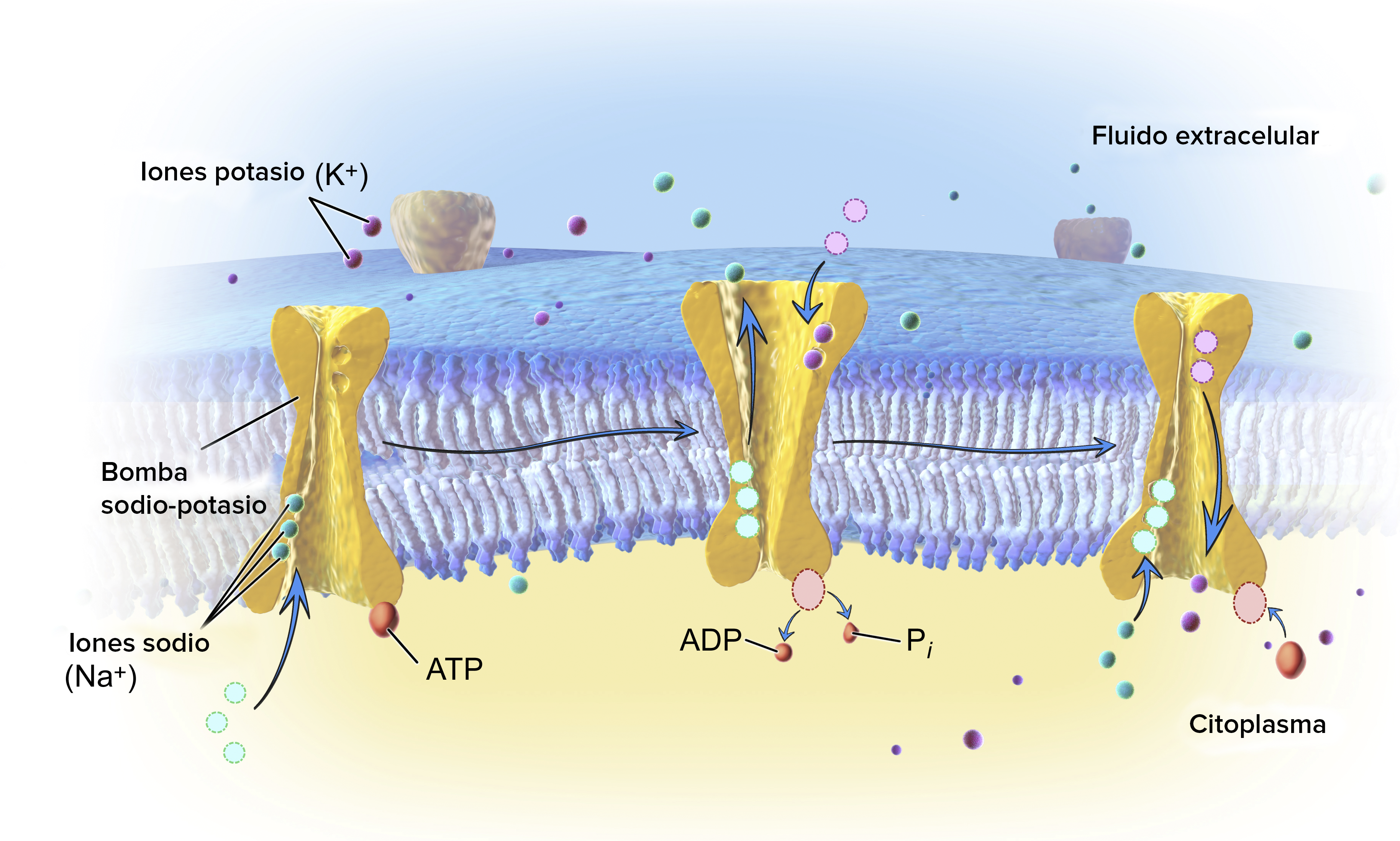
Por las razones dadas, desde un punto de vista funcional, resulta útil reducir las subdivisiones anteriores a dos: medio intracelular y medio interno. Ambos medios se encuentran separados por la membrana celular que, dada su carácter semipermeable, ejerce de barrera para con ciertos solutos (Na+, Cl–, sustancias orgánicas) pero no para con el K+ ni para con el agua. Y por otro lado, el medio interno y el medio exterior también se encuentran separados por el tegumento, que normalmente hace función de barrera.
Los animales han de preservar su contenido hídrico dentro de ciertos márgenes, porque de ese contenido depende la existencia de los fluidos corporales y que esos fluidos tengan características compatibles con las funciones animales. Al fin y al cabo, todas las estructuras internas se hallan bañadas por fluidos, que son el ambiente inmediato de células, orgánulos celulares y moléculas. Su composición es el contexto en el que funcionan aquellas. Las concentraciones de los iones a los que nos referiremos enseguida dependen del volumen de los fluidos en los que se encuentran disueltos. Y además, de ese volumen depende el de las células y tejidos.
Los animales también han de mantener en un rango restringido de valores la concentración intracelular de los iones inorgánicos. El K+, principal catión intracelular, ha de mantener una concentración relativamente alta, pues así lo requiere la estabilidad e integridad funcional de las proteínas. Del mismo modo, han de mantener a niveles reducidos la concentración de Na+ y Cl– dentro de las células, porque incluso concentraciones moderadas de esos iones desestabilizan las proteínas celulares. Además, las concentraciones intra y extracelulares de esos tres iones puede determinar (dependiendo de la permeabilidad relativa de la membrana celular para con cada uno de ellos) la diferencia de potencial eléctrico entre los lados interno y externo de la membrana, lo que resulta crucial desde el punto de vista de la transmisión del impulso nervioso en las neuronas y de la contracción muscular.
Por todo lo apuntado hasta aquí, desde el punto de vista de los equilibrios que han de mantener los animales en materia de contenido hídrico y concentración interna de solutos, la capacidad para ejercer un cierto control sobre los movimientos de agua y sales a través de esas barreras es esencial en la mayor parte de las especies. Aunque ese control se ejerce de forma diferente en las dos barreras mencionadas (membrana celular y tegumento corporal) dependiendo del tipo de relación que cada especie mantiene con el medio en el que se encuentra.
Los animales, como es sabido, ocupan medios acuáticos y el medio terrestre. En lo relativo a los medios acuáticos, estos pueden tener muy diferentes concentraciones salinas. No obstante, la división fundamental es la que existe entre el agua de mar y las aguas dulces. El agua marina tiene una concentración osmótica de 1000 mOsm (miliosmolar), que corresponde a unos 35 g kg-1 (g de sales por kg de agua). Las aguas dulces varían dentro de un rango relativamente estrecho, de entre 0,15 y 15 mOsm; en términos de masas, su concentración de sales se encuentra entre 0,1 y 0,2 g kg-1, aunque se consideran aguas dulces todas aquellas cuya concentración gravimétrica es inferior a 0,5 g kg-1. Los iones principales del agua de mar son Na+ (470 mM), Cl– (548 mM), Mg+2 (54 mM), SO4-2 (28 mM), Ca+2 (10 mM) y K+ (10 mM). Las concentraciones de esas sustancias en aguas dulces, aunque muy bajas, pueden variar considerablemente entre unas masas de aguas y otras. No obstante, las siguientes pueden considerarse típicas del agua dulce: Na+ (0,35 mM), Cl– (0,23 mM), Mg+2 (0,21 mM), SO4-2 (0,19 mM), Ca+2 (0,75 mM) y K+ (0,08 mM).
Por otra parte, también hay importantes volúmenes de agua cuya concentración salina se encuentra entre la del mar y la del agua dulce: son las aguas salobres. Son relativamente normales en zonas costeras en las que el aporte de agua dulce continental diluye el agua marina, especialmente en los estuarios. Y también hay aguas hipersalinas, normalmente en lagos que experimentan una fuerte evaporación; son las aguas cuya concentración supera claramente la del agua de mar.

En lo que se refiere a los medios terrestres, no existen medios que puedan ser diferenciados de forma tan nítida como las aguas saladas y las aguas dulces. La variable relevante en este caso es la presión parcial de vapor de agua en la atmósfera, porque esa variable, como veremos, afecta directamente a la pérdida de agua de un animal por evaporación. La presión de vapor de agua es la porción de la presión atmosférica que es debida al gas vapor de agua y tiene la peculiaridad, por comparación con otros gases, de que tiene un valor límite que no puede superar. Ese valor, que es la presión de saturación, varía con la temperatura: cuanto más alta es esta, mayor es la presión de vapor de saturación: a 0ºC, la presión de saturación del vapor de agua es 4,6 mmHg (0,61 kPa) y a 40ºC es 55,3 mmHg (7,37 kPa).
El agua que puede perder por evaporación un animal depende (1) de la presión parcial de vapor de agua en la atmósfera en la que se encuentra y de lo próxima o alejada que se encuentre esa presión de la de saturación (cuanto mayor sea la diferencia entre la presión de saturación y la presión parcial de vapor de agua, mayor será la pérdida), (2) de la renovación del aire en la proximidad del tegumento, (3) de la temperatura corporal del animal (por razones que no detallaremos aquí), (4) del grosor de la barrera que separa de la atmósfera el fluido corporal que se evapora y (5) de la permeabilidad del tegumento para con el agua. Por lo tanto, los factores ambientales que más inciden en la pérdida potencial de agua por evaporación son la presión parcial de vapor del agua, el viento, la temperatura del ambiente (porque de ella depende la presión de saturación) y la temperatura del animal. El agua se puede perder por otras vías, pero no nos ocuparemos aquí de ellas pues no guardan relación directa con los factores ambientales, que son los que ahora nos interesan. Por lo mismo, tampoco nos ocuparemos de los mecanismos para adquirir agua.
Los animales pueden desarrollar tres tipos de regulación: osmótica, iónica y de volumen. La regulación osmótica (osmorregulación) consiste en el mantenimiento en niveles constantes o casi constantes la concentración osmótica del medio interno. Cuando se define de esta forma la osmorregulación, normalmente se asume que tal capacidad se ejercita en el contexto de variaciones ambientales en la concentración osmótica del medio externo. Pero eso no tiene por qué ser así. También cabe hablar de osmorregulación en aquellos casos en que los animales mantienen concentraciones osmóticas internas constantes (o casi constantes) y diferentes de la del medio externo. Incluso si nos referimos a un animal terrestre que vive en un medio no acuático y que, por tanto, no puede ser caracterizado en términos de concentración osmótica, también podemos considerarlo osmorregulador si es capaz de mantener constante la concentración osmótica del medio interno en un contexto, por ejemplo, de variaciones en la disponibilidad de agua o bajo regímenes alimenticios de variables contenidos salinos.
Los animales que no regulan la concentración osmótica de su medio interno son osmoconformadores, lo que quiere decir que, si ocupan medios acuáticos la concentración osmótica de su medio interno se asemeja a la del medio externo y que si se trata de animales terrestres, varía en función de las condiciones ambientales.
La regulación iónica, por su parte, consiste en la capacidad para mantener constante la concentración de iones inorgánicos en el medio interno. En este caso no cabe hablar de una capacidad de regulación iónica general, sino que tal capacidad siempre se refiere a un ión concreto puesto que cada ión cuya concentración está regulada está sujeto a procesos específicos. También en relación con los iones inorgánicos recurrimos al término conformador para denominar al animal que no regula la composición de alguno de ellos; como en el caso de la regulación, también aquí la incapacidad para regular la concentración de un ión es específica para ese ión.
Nos interesa también la capacidad –o su ausencia- para regular el volumen del medio interno. Si un animal dispone de tal capacidad diremos que es un regulador del volumen, y que es conformador del volumen si carece de ella.
Para terminar, interesa valorar qué consecuencias tiene para el medio intracelular que un animal sea o no capaz de regular las magnitudes anteriores. Dado que las membranas celulares son permeables al agua, la concentración osmótica del medio intracelular es la misma que la de su medio interno; por lo tanto, un animal que regule este, mantendrá aquel igualmente regulado. Si, por el contrario, no regulase osmóticamente su medio interno, el medio intracelular experimentará una variación tendente a igualar su concentración con la de aquel, y dado que los iones inorgánicos han de ser mantenidos en un rango estrecho de concentraciones, ello podría provocar cambios en el volumen de las células debidas a flujo osmótico de agua.
En lo que a la concentración de iones inorgánicos se refiere, es importante que los ya señalados Na+, Cl– y K+ se mantengan dentro de un margen relativamente estrecho de concentraciones propias de los medios interno e intracelular. Las variaciones que se produzcan en el medio interno no deberían ser importantes ni incidir en lo que ocurra dentro de las células. De lo contrario, y como ya se ha señalado, la estructura de las proteínas (enzimas, principalmente) y, por ende, su integridad funcional se vería amenazada.
En lo que al volumen se refiere, la capacidad o incapacidad para regular el volumen del medio interno no debería afectar al volumen del medio intracelular si no se producen cambios en la osmolaridad. No obstante, en el caso de que un cambio en el volumen del medio interno ocurriese a la vez que una variación en su concentración osmótica, el volumen celular podría verse afectado como ya se ha dicho antes. No obstante, muchos animales han desarrollado la capacidad de realizar ajustes de la concentración intracelular de osmolitos orgánicos gracias a los cuales pueden mantener constante su volumen celular (regularlo) aunque cambie la concentración osmótica fuera (y dentro) de las células. Gracias a esa capacidad, a la que nos referimos aquí, se evita el flujo osmótico de agua hacia o desde el interior celular y el volumen permanece constante. A esa capacidad se la denomina regulación isosmótica del volumen celular.
Sobre el autor: Juan Ignacio Pérez (@Uhandrea) es catedrático de Fisiología y coordinador de la Cátedra de Cultura Científica de la UPV/EHU

Animales en equilibrio osmótico: invertebrados marinos y peces bruja – Cuaderno de Cultura Científica
[…] composición iónica similar, aunque no idéntica, a la del agua de mar. Por las razones que dimos aquí, el medio intracelular, sin embargo, tiene una composición de solutos muy diferente ya que una […]
Economía del agua en animales terrestres – Cuaderno de Cultura Científica
[…] vimos aquí, la pérdida de agua de los animales por evaporación depende de un conjunto de factores: (1) […]
Zacarías Blancarte
Un enfoque sistematico y de un nivel accesible.Uno de los pocas exposiciones utiles de la biologia marina.