La función renal en vertebrados

La mayor parte de lo que sabemos sobre el funcionamiento de los riñones de vertebrados procede de estudios hechos con anfibios y con mamíferos. Los anfibios resultan ser un modelo animal muy útil a estos efectos, y lo que sabemos del funcionamiento de su riñón es generalizable a la mayoría de vertebrados y, en concreto, a los peces de agua dulce y a los reptiles, exceptuando en parte a las aves.
En la nefrona de la mayor parte de grupos de vertebrados se diferencian los siguientes elementos: (1) cápsula de Bowman, (2) túbulo contorneado proximal, (3) segmento intermedio, (4) túbulo contorneado distal, y (5) túbulo colector. Los segmentos llamados “túbulo contorneado” se caracterizan por que describen múltiples curvas en su trayectoria; a eso se refiere el término “contorneado”. En inglés se dice convoluted, que puede traducirse también como enmarañado, retorcido o complicado; el término “contorneado” es claramente más compasivo. El segmento intermedio es bastante recto y de muy pequeño diámetro, y el túbulo colector es también relativamente recto, pero de mayor grosor que el intermedio. Los segmentos desempeñan diferentes funciones, aunque también pueden cambiar dentro de cada segmento. La longitud de la nefrona viene a ser de 1 cm aproximadamente, aunque el grosor es microscópico en todo su recorrido. El túbulo colector desemboca, junto con muchos otros túbulos, en el ducto o tubo colector, y los ductos colectores desembocan en el uréter, que es el canal que conduce la orina a la vejiga.
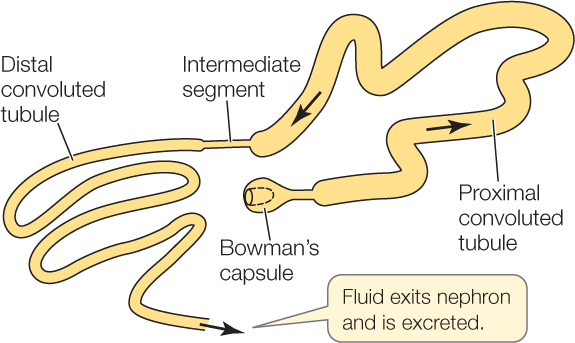
Como vimos en la anotación anterior, lo normal es que una parte muy importante de la orina primaria no sea expulsada al exterior, sino que sea reabsorbida a lo largo de la nefrona, y esa reabsorción afecta tanto al agua como a las sales y otras sustancias que conviene retener. En el caso de los anfibios también es ese el caso, aunque no suele ser agua lo que les falta. De hecho, para la mayor parte de los animales que viven en agua dulce el problema consiste en retener las sales a la vez que expulsan la máxima cantidad de agua posible.
A lo largo del túbulo contorneado proximal se reabsorbe activamente Na+, y lo normal es que el Cl– lo acompañe al moverse a favor del gradiente eléctrico generado por el transporte del catión. La concentración osmótica a lo largo de ese segmento apenas varía, porque su epitelio es permeable al agua, de manera que la reabsorción de NaCl se ve acompañada por una reabsorción de agua de similar magnitud. En los anfibios estudiados, entre un 20% y un 40% de la orina primaria es reabsorbida en este primer segmento de la nefrona. Además de NaCl y agua, en el túbulo proximal se reabsorbe glucosa y aminoácidos.
La reabsorción de NaCl prosigue en el túbulo contorneado distal, aunque en este caso, la recuperación de sal puede ir acompañada de reabsorción de agua, o no, dependiendo de las necesidades que impone el mantenimiento del balance hídrico. En otras palabras, el volumen final de orina que se expulsa depende de la medida en que el agua acompaña al NaCl cuando es reabsorbido este en el túbulo distal; cuanto menor es la reabsorción de agua, mayor es la producción de orina final (mayor es la diuresis) y cuanto mayor es el volumen que se reabsorbe, menor es la diuresis. En algunas especies de anfibios la vejiga también interviene en la formación de orina de forma similar a como lo hace el túbulo contorneado distal.
La permeabilidad para con el agua del epitelio del túbulo contorneado distal está controlada por la hormona antidiurética (ADH) o vasotocina1, una hormona peptídica producida por la neurohipófisis; y ese control se ejerce a través de la inserción de más o menos aquaporinas en la membrana apical de las células epiteliales. Cuando los niveles circulantes de esta neurohormona son bajos, la permeabilidad del epitelio es baja (menor inserción de aquaporinas), de manera que la reabsorción de NaCl por las células epiteliales del túbulo distal no es acompañada por una reabsorción equivalente de agua. Como consecuencia, se recupera poca agua, la orina producida es abundante y además muy diluida, esto es, su concentración de sales es muy baja. Lo contrario ocurre cuando la concentración circulante de vasotocina es alta: hay una intensa reabsorción de agua en el túbulo distal (en su parte más próxima al túbulo colector, principalmente) y, en consecuencia, la orina es escasa y su concentración salina es más alta. Estos son, precisamente, los procesos mediante los que se efectúa la regulación del volumen de los líquidos coporales y de la concentración osmótica y salina que realizan muchos animales y que vimos en algunas anotaciones de esta serie.
En anfibios, lagartos, aves y otros reptiles la hormona antidiurética no solo aumenta la permeabilidad al agua de ciertas zonas del túbulo contorneado distal, sino que también disminuye la tasa de filtración glomerular, mediante una reducción en el número de nefronas que filtran plasma sanguíneo. Esa reducción se produce provocando vasoconstricción en los vasos que llevan sangre al glomérulo (aferentes). Este mecanismo complementa el descrito anteriormente y ambos contribuyen a retener agua y limitar el volumen de orina que es expulsada al exterior. Y en algunas especies de anfibios la ADH eleva, además, la reabsorción activa de NaCl, favoreciendo también de este modo la reabsorción de agua.
En el mecanismo que desencadena la elevación de la concentración sanguínea de vasotocina intervienen osmorreceptores hipotalámicos (que detectan una subida de la concentración osmótica) y barorreceptores o receptores de estiramiento localizados en ciertos vasos sanguíneos (que detectan cambios en la presión sanguínea). Así, ante un aumento en la concentración osmótica o una reducción en la presión sanguínea, la neurohipófisis responde secretando ADH y elevando así su concentración sanguínea para surtir los efectos descritos y restaurar el volumen de sangre y la concentración osmótica adecuada.
Los riñones de los teleósteos marinos presentan algunas características que los diferencian de los de agua dulce y de los anfibios. Lo normal es que las nefronas de los peces marinos carezcan del túbulo contorneado distal. Puesto que estos peces proceden, de hecho, de antecesores dulceacuícolas, se cree que esa ausencia es secundaria y se atribuye al hecho de no necesitar producir grandes volúmenes de orina diluida. Los peces marinos son reguladores hiposmóticos y el riesgo principal que afrontan es el de la desecación. Además, y por la misma razón, estos peces tienden a tener pocas nefronas y glomérulos pequeños, pues de esa forma limitan la formación de orina primaria, reduciendo significativamente la pérdida de agua por esa vía. En coherencia con esas características renales, no toda la orina primaria la producen por ultrafiltración. De hecho, parte de la orina la generan por secreción activa de iones (Na+, Cl–, Mg2+ y SO42-), a los que acompaña el agua, a la luz del túbulo contorneado proximal.
La reducción de la contribución de la ultrafiltración a la formación de orina primaria alcanza su máxima expresión en un conjunto de especies (se conocen una treintena) de peces marinos cuyas nefronas carecen de glomérulo, son aglomerulares. Se trata de un rasgo que ha aparecido en tres ocasiones diferentes en el curso de la evolución.
Las aves, a diferencia del resto de reptiles, tienen dos tipos de nefronas. Unas son similares a las descritas en esta anotación, pero las otras se asemejan a las de los mamíferos (que veremos más adelante). Entre un 10% y un 30% de las nefronas aviares son de este segundo tipo y se caracterizan por tener lo que se conoce como “asa de Henle”, un segmento que en mamíferos cumple un papel de importancia crucial. En el riñón de las aves las nefronas de tipo anfibio (o reptiliano) se disponen en la corteza renal (parte más exterior), mientras que las de tipo mamífero se disponen en la médula, la zona más interior y más próxima al uréter. Lógicamente, las características funcionales de las nefronas de tipo mamífero se asemejan a las de los mamíferos, por lo que pueden producir una orina de mayor concentración osmótica que la sangre, aunque no en una medida equivalente a la de aquéllas.
Por último, cabe señalar que el uréter de las aves no descarga en una vejiga o directamente al exterior, sino que lo hace en la cloaca, de forma similar a como vimos con los túbulos de Malpigio y el recto en insectos. Y esta no es la única semejanza entre aves e insectos, ya que las aves también eliminan sus restos nitrogenados en forma de ácido úrico, que aparece en la nefrona por filtración o, incluso, por secreción activa. También las aves eliminan sus restos nitrogenados en forma semisólida, lo que ayuda de forma importante al ahorro de agua. En próximas anotaciones presentaremos la nefrona de mamíferos.
1 La arginina vasotocina (AVT) es la equivalente en vertebrados no mamíferos a la arginina vasopresina (AVP) de mamíferos.
Sobre el autor: Juan Ignacio Pérez (@Uhandrea) es catedrático de Fisiología y coordinador de la Cátedra de Cultura Científica de la UPV/EHU
Hitos en la red #228 – Enlaces Covalentes
[…] La función renal en vertebrados […]