Túbulos de Malpigio y recto, el sistema excretor de los insectos

Los túbulos de Malpigio son ductos ciegos cuyo epitelio tiene el grosor de una célula. Se encuentran suspendidos en la hemolinfa y desembocan en el intestino posterior, al que vierten sus productos. Se disponen por pares a ambos lados del cuerpo, pudiéndose proyectar unos hacia adelante y otros hacia atrás. El número de túbulos varía con las especies: Drosophila melanogaster, la conocida mosca de la fruta, tiene dos pares, pero la langosta del desierto tiene centenares.
La orina no se forma en los túbulos de Malpigio por filtración, como ocurre en protonefridios y metanefridios, sino que surge a partir de la secreción activa de K+ -y en ocasiones también de Na+– hacia la luz del túbulo. En ese transporte no participa la muy común ATPasa de Na+/K+, sino una ATPasa V, no tan común pero muy antigua. Al parecer, la ATPasa V genera un gradiente de protones entre la luz del túbulo (alta concentración) y la hemolinfa (baja concentración), de manera que el movimiento de protones desde el interior del túbulo hacia el medio interno impulsa el movimiento en sentido contrario de K+ (y Na+), mediante el concurso de transportadores CPA (antiporters catión-protón) localizados en determinadas células epiteliales de los túbulos (en los dípteros son las denominadas células principales). Este dispositivo permite transportar ciertas sustancias contra gradiente gracias a la energía que aporta una sustancia que se mueve a favor de su gradiente de concentración, protones en este caso.
Al acumularse Na+ y K+ en la luz del túbulo, su contenido se vuelve positivo, lo que impulsa el movimiento de iones Cl– a favor de gradiente eléctrico. Se cree que el Cl– atraviesa el epitelio por canales ClC (canales de Cl– activados por Cl–) de las células estrelladas del epitelio tubular, en unas especies, y a través de vías paracelulares, en otras. La acumulación de KCl y en ocasiones de NaCl genera un gradiente de concentración osmótica que impulsa el movimiento de agua desde la hemolinfa hacia la luz del túbulo. Ese movimiento también se puede producir atravesando las células epiteliales, mediante acuaporinas, o a través de los espacios paracelulares. De esa forma surge la orina primaria.
A partir de su generación, esa orina primaria ve modificada su composición debido a la secreción activa –mediante transportadores- o el movimiento pasivo –a través de canales proteicos- de ciertas sustancias. El ácido úrico, que es la molécula de excreción de restos nitrogenados en los insectos, es secretada de forma activa por transportadores específicos. Además de canales y transportadores, en el epitelio de los túbulos de Malpigio hay enzimas detoxificadoras, que están relacionadas con las del hígado de vertebrados, cuyo cometido es, lógicamente, el de eliminar sustancias tóxicas.
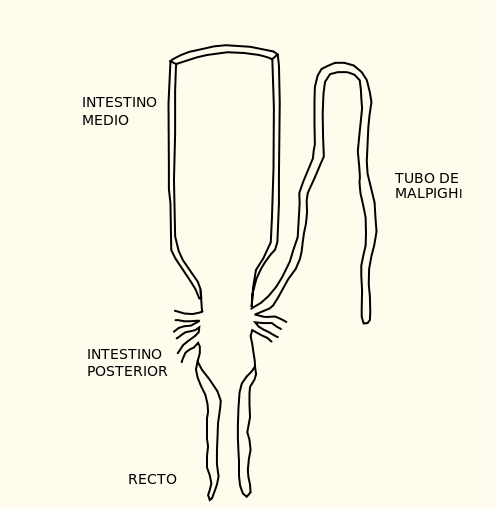
El fluido formado en los túbulos llega al intestino posterior con una concentración osmótica similar a la de la hemolinfa, aunque con una composición muy diferente. Ciertas células especializadas del recto pueden seguir modificando las características del fluido antes tubular y ahora rectal. Su papel es especialmente necesario en condiciones de baja disponibilidad de agua. En esos casos se puede producir una intensa reabsorción de agua, en la que está implicado también el KCl, cuya reabsorción en el recto permite que se produzca el movimiento de agua desde la luz rectal a la hemolinfa. Esa reabsorción de agua puede dar lugar a una fuerte elevación de la concentración osmótica, lo que unido a la secreción activa de protones hace que el ácido úrico llega a precipitar, de manera que es expulsado al exterior en forma de uratos sólidos o semisólidos.
Un aspecto especialmente reseñable del funcionamiento del sistema excretor de insectos es que, como veremos que ocurre en aves y en mamíferos, este también es capaz de producir una orina de mayor concentración osmótica que el medio interno. Y esa producción conlleva, por paradójico que resulte, la reabsorción de agua en el recto incluso en contra de fuertes gradientes de concentración osmótica. Esa capacidad paradójica se basa en tres mecanismos alternativos.
En ciertas especies son las denominadas papilas rectales del intestino posterior las responsables de la ejecución de tan asombrosa tarea. En ellas, la disposición microanatómica de sus elementos constituyentes (el epitelio, los espacios intercelulares y los denominados canales infundibulares) permite mantener una especie de flujo circular de KCl (alimentado mediante transporte activo), de manera que el movimiento de la sal “arrastra” agua (provoca su salida de la luz rectal) y posteriormente, una vez generado el flujo, aquella (o sea, la sal) se recupera y se mantiene en el flujo circular conforme el fluido, con una baja concentración salina, retorna a la hemolinfa.
Un segundo dispositivo, propio de especies que viven en ambientes muy secos, es el complejo criptonefridial. Ese complejo se basa en una gran proximidad física entre la zona ciega del túbulo de Malpigio y el epitelio rectal, y en el transporte activo de KCl y NaCl desde la luz del recto hasta la del túbulo. El epitelio del intestino posterior permite el paso del agua, de manera que así se reabsorbe desde el fluido rectal hasta la hemolinfa, pero el agua no puede atravesar el epitelio tubular de la zona implicada en este proceso (porque es impermeable), por lo que se genera un fortísimo gradiente de concentración osmótica entre el túbulo y el recto, debido a su proximidad física. Ese dispositivo actúa como una bomba osmótica de succión de agua.
El tercer mecanismo es característico de las pocas especies que viven en aguas salinas. Y se basa en la secreción activa de sales al fluido rectal, lo que permite, mediante transporte activo una vez más, elevar la concentración de la orina contra gradiente osmótico.
En las especies que utilizan néctar como alimento o en los insectos acuáticos el agua no es un factor limitante, aunque su abundancia podría ser un problema. En esos casos los insectos eliminan abundante orina hipotónica, de menor presión osmótica que la hemolinfa, para lo que el epitelio rectal recupera activamente las sales que han servido para la formación de la orina primaria, sin que ello lleve emparejada reabsorción de agua. Hay también, como es sabido, insectos ectoparásitos, como mosquitos o chinches besuconas, que se alimentan de la sangre de sus hospedadores. Estos llegan a multiplicar por diez la masa corporal gracias a la sangre recién ingerida. Pues bien, estas especies empiezan a producir y a eliminar orina rápidamente, antes incluso de que hayan terminado de comer. Es una orina isosmótica, normalmente, por lo que su eliminación no conlleva especiales complicaciones, pero ha de cursar a gran velocidad, razón por la cual las chinches besuconas llegan a multiplicar por mil la velocidad a la que producen y expulsan la orina.
El sistema formado por los túbulos de Malpigio y el recto es, como hemos visto, muy flexible, puesto que puede dar satisfacción a necesidades muy dispares en lo que al balance hídrico se refiere. Esa flexibilidad se basa en el recurso al transporte activo de sales y a unas disposiciones microanatómicas complejas que permiten recuperar agua en condiciones en que el resto de especies de invertebrados no pueden hacerlo. Es posible que esa gran flexibilidad haya sido una de las claves del gran éxito de este grupo, al que pertenece el mayor número de especies animales existentes.
Sobre el autor: Juan Ignacio Pérez (@Uhandrea) es catedrático de Fisiología y coordinador de la Cátedra de Cultura Científica de la UPV/EHU