La función del asa de Henle en el riñón de mamíferos
Si se compara con la de anfibios, reptiles (no aviares) o peces de agua dulce, la nefrona de mamíferos tiene un elemento del que carecen las anteriores. Ese elemento es un segmento que se encuentra entre el túbulo proximal y el túbulo distal y que tiene la forma de una horquilla para el pelo. Se llama “asa de Henle”. Otra diferencia es que las asas de Henle y los ductos colectores se disponen en batería, lo que confiere a los riñones de mamíferos una macroestructura bien definida de la que carecen los riñones del resto de los grupos, salvo los de las aves. Y son precisamente estas dos características distintivas las que permiten al riñón de mamíferos producir una orina de mayor concentración osmótica que la de la sangre. Para hacernos una idea de la importancia de este rasgo, conviene recordar que peces, anfibios, lagartos, serpientes, cocodrilos y tortugas carecen de esa facultad, y que en el dominio animal tan solo insectos, aves y mamíferos han desarrollado la capacidad de producir orina hiperosmótica con relación a la sangre. Gracias a ella estos grupos han gozado de grandes posibilidades para colonizar una gran variedad de medios, incluyendo algunos tan exigentes como los desiertos más secos del planeta.
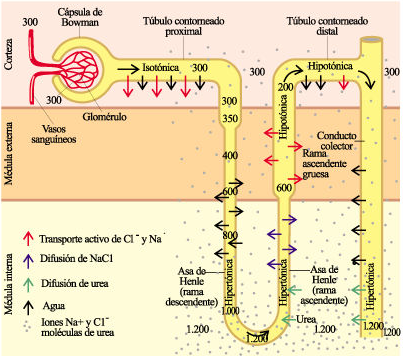
La cápsula de Bowman y su glomérulo se disponen en la zona exterior (corteza) del riñón, así como el túbulo proximal que emerge de la cápsula de Bowman. El asa de Henle tiene una rama descendente, que se dirige a la médula renal, y una ascendente, que vuelve hacia la corteza. Las nefronas de estos riñones pueden tener asas de Henle de diferentes dimensiones. En las que tienen el asa de Henle más larga la primera parte de su rama descendente es un segmento relativamente grueso, que da paso enseguida a otro más fino. También en la rama ascendente se diferencian un segmento fino, que es continuación del de la rama descendente, y uno grueso (ambos segmentos de similares dimensiones) hasta que llega al túbulo distal, que se dispone junto a la cápsula de Bowman en la corteza. En las nefronas de asa más corta esta penetra muy poco en la médula renal y el segmento fino es de longitud muy reducida. Por lo tanto, túbulos contorneados proximal y distal y cápsula de Bowman se encuentran en la corteza renal, mientras que el asa de Henle y los ductos colectores, que discurren paralelos unos a otros, se proyectan hacia el interior, en la médula renal. Las nefronas de asa larga tienen la cápsula de Bowman en una posición más próxima a la médula renal, aunque siempre en la corteza, mientras que las de asa más corta tienden a estar más alejadas de la médula y próximas a la superficie renal.
El segmento ascendente grueso de cada nefrona, al convertirse en túbulo distal, pasa junto a la cápsula de Bowman de su misma nefrona. Y en ese punto, en la pared del segmento aparecen un conjunto de células especializadas denominado mácula densa. En posición adyacente, pero en el endotelio de la arteriola aferente se encuentra otro conjunto de células especializadas, denominadas células yuxtaglomerulares o granulares. El conjunto conforma una estructura que se denomina aparato yuxtaglomerular. Las células yuxtaglomerulares secretan renina, una sustancia que, a su vez, controla la secreción de otra hormona, la aldosterona, que promueve la reabsorción renal de sodio.
A medidados del siglo XX los estudios de anatomía comparada mostraron que los riñones con una médula más gruesa eran capaces, por regla general, de producir una orina más concentrada osmóticamente, y solían pertenecer a animales propios de medios secos. A partir de esos elementos, se empezó a desentrañar el funcionamiento del riñón de mamíferos y el papel que en ese funcionamiento jugaba el asa de Henle.

La orina que llega al ducto colector desde la nefrona tiene una concentración de solutos inorgánicos (Na+, Cl–, K+ y SO42-, principalmente) inferior a la de la sangre. Sin embargo, dependiendo de las necesidades fisiológicas del organismo y, más concretamente, de si se encuentra en una situación de antidiuresis, conforme la orina discurre a lo largo del ducto colector, la concentración de los solutos inorgánicos se eleva hasta alcanzar valores muy superiores a los de la sangre. Eso ocurre porque el agua sale del ducto colector a favor de gradiente osmótico, gradiente que es debido a la alta concentración de NaCl en el fluido intersticial de la médula; además, esa concentración es mayor cuanto más penetra el tubo colector en la médula renal. Para que el gradiente osmótico dé lugar a la reabsorción de agua, es preciso que el epitelio del ducto colector sea permeable al agua, que es lo que ocurre cuando se dan condiciones fisiológicas de antidiuresis. Los solutos inorgánicos de la orina antes citados, por otra parte, no pueden atravesar el epitelio del ducto colector, de lo contrario el gradiente osmótico se atenuaría y el agua no saldría.
El gradiente de concentración de NaCl es debido a los procesos que ocurren en el asa de Henle. El epitelio de su rama ascendente transporta activamente NaCl desde la luz del túbulo hasta el fluido intersticial de la médula. Y además es un epitelio impermeable al agua. De esa forma, disminuye la concentración de la sal en la orina conforme esta transita por la rama ascendente del asa de Henle; y a la vez, la concentración de la sal es alta en el fluido intersticial. Por otra parte, aunque las características de la rama descendente difieren de unas especies a otras, lo normal es que de una forma o de otra, la concentración osmótica de la orina que se desplaza por su interior se equilibre con la del fluido intersticial.

Ese mecanismo genera una diferencia de concentración osmótica entre las dos ramas del asa de Henle que puede llegar a ser de 200 mOsm en su parte superior, cerca de la corteza. Sin embargo, el trasiego de NaCl que se produce a lo largo de toda la rama ascendente da lugar a que la concentración osmótica sea muy alta (alrededor, por ejemplo, de 700 mOsm o más) en la zona del codo, mientras que en la zona de la corteza llega a ser muy baja, (alrededor de 200 mOsm o menos), de manera que la diferencia entre las dos zonas, la médula interior y la corteza, llega a ser de 500 o 600 mOsm, bastante mayor que la que hay entre las dos ramas. A ese mecanismo se le denomina multiplicador contracorriente, porque multiplica el gradiente osmótico valiéndose de dos conductos por los que circula un fluido en sentidos opuestos y en íntima proximidad. En este esquema es muy importante el hecho de que todas las nefronas se encuentren dispuestas en batería, porque es la actividad conjunta de todas ellas la que da lugar a que el fluido intersticial de la médula interior tenga una alta concentración osmótica, mientras que en la corteza sea mucho más baja. Ha de tenerse en cuenta que son miles las nefronas que forman un riñón de mamífero: los de las ratas tienen alrededor de 30.000 cada uno, los de los perros tienen alrededor de 400.000, y los de los seres humanos entre 400.000 y 1.200.000.
Me he referido antes al grosor de la médula renal, señalando que las especies de mamíferos con una médula más gruesa producen una orina más concentrada y habitan entornos más secos. La razón de esa correlación es que médulas de mayor grosor relativo (con respecto al grosor renal total o al tamaño del animal) pertenecen a riñones en los que hay una mayor proporción de nefronas de asas de Henle largas. Los roedores que habitan zonas séricas tienen riñones de un gran grosor medular y todas sus nefronas son de asa larga. En el otro extremo están los castores, por ejemplo, que viven rodeados de agua; sus riñones no tienen nefronas de asa larga, todas son de asa corta.
Sobre el autor: Juan Ignacio Pérez (@Uhandrea) es catedrático de Fisiología y coordinador de la Cátedra de Cultura Científica de la UPV/EHU