La regulación de la diuresis en mamíferos
Al contrario que otros vertebrados, los mamíferos (salvo algunas especies, como los dromedarios) no ajustan la producción de orina modificando la tasa de filtración glomerular (GFR). De hecho mantienen esta relativamente constante y modifican la fracción de la orina primaria que es reabsorbida antes de su evacuación al exterior para regular el volumen y la concentración osmótica de los fluidos corporales.
Un porcentaje relativamente alto de la orina primaria –entre un 60% y 80% del volumen- es reabsorbido desde el túbulo contorneado proximal, para lo cual se reabsorben activamente porcentajes similares de NaCl. Además de la sal y el agua, glucosa, aminoácidos y bicarbonato son también reabsorbidos en ese segmento, estos prácticamente en su totalidad.
La reabsorción de Na+ se produce gracias a la actividad de una ATPasa de Na+-K+ de la membrana basolateral del epitelio (la que separa el líquido intersticial del interior celular). Ese transporte genera un gradiente electroquímico en la membrana apical (la que separa el interior celular de la luz del túbulo) que favorece la entrada de sodio desde el fluido tubular. La reabsorción de agua se produce a través de aquaporinas estructurales, que se hallan siempre presentes en esas células (su presencia apenas depende de factores externos).
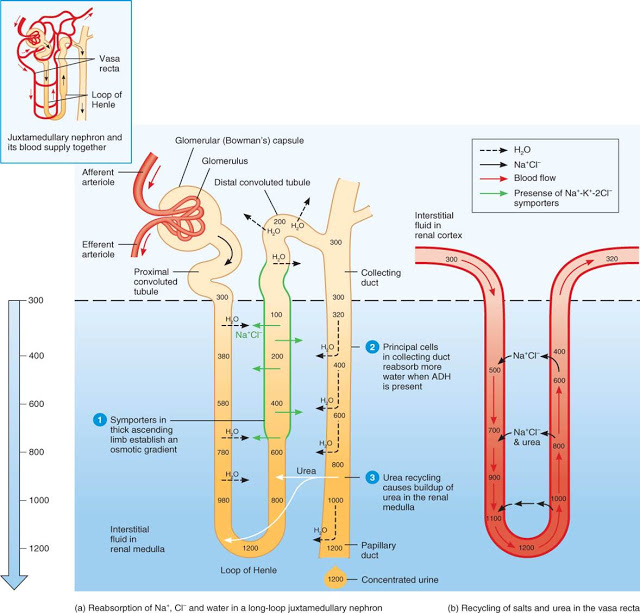
Tras pasar por el túbulo proximal, la orina penetra en el asa de Henle. Lo hace con una concentración osmótica de unos 300 mOsm, pero tras su recorrido por ese segmento, puede alcanzar el túbulo distal con una concentración inferior a la anterior (puede llegar a ser de 100 mOsm). Como vimos, esa diferencia es debida al transporte activo de NaCl que tiene lugar en la rama ascendente, que lo retira de la orina, disminuyendo en ella su concentración, a la vez que aumenta la del líquido intersticial.
Tras el asa de Henle la orina accede al túbulo contorneado distal. El epitelio de ese segmento transporta NaCl activamente de la luz del túbulo al especio intersticial. Y aunque no es impermeable, es poco permeable al agua.
Lo que ocurre a continuación en el tubo colector depende de cuáles son las necesidades hídricas del organismo. Si necesita retener agua, el organismo se encuentra en estado de antidiuresis y el riñón producirá un volumen de orina muy limitado. Eso es consecuencia de una intensa reabsorción de agua en el tubo colector, reabsorción que se produce gracias a la presencia en la membrana apical de las células de la pared del tubo de numerosas moléculas de una forma de aquaporina (AQP-2) específica de ese epitelio. Recordemos que debido al transporte activo de NaCl que tiene lugar en la rama ascendente del asa de Henle y al equilibrio iónico y osmótico que se establece entre su rama descendente y el fluido intersticial de la médula renal, la concentración osmótica de ese fluido en el interior medular es muy alta. Por ello, la presencia de numerosos poros en las membranas apicales de las células de la pared del tubo colector permiten que el agua pase con gran facilidad de un fluido que se encuentra originariamente a una concentración osmótica muy baja (la orina que llega del túbulo distal) a otro con la concentración osmótica muy alta (el fluido intersticial medular). Y eso ocurre hasta que ambas concentraciones osmóticas se igualan, para lo que debe pasar un volumen muy importante de agua. Además, a lo largo del tubo colector se sigue reabsorbiendo activamente NaCl, lo que favorece aún más este proceso. Como consecuencia de esa reabsorción, la orina final puede llegar a representar tan solo un 1% del volumen de plasma filtrado en el glomérulo y alcanzar una concentración osmótica de 1200 mOsm, que es cuatro veces más alta que la del plasma. Pero en mamíferos con grandes restricciones hídricas esa concentración puede llegar a multiplicarse por diez o más. Producen mínimas cantidades de orina y evitan así perder agua por esa vía.

Ese estado de antidiuresis a que me he referido en el párrafo anterior es el que se produce cuando hay una alta concentración sanguínea de la hormona antidiurética (ADH), que en mamíferos es la arginina vasopresina (AVP). Es ella la responsable de la presencia en las células de la pared del ducto de numerosas unidades de aquaporinas AQP-21. Por ello, cuando las condiciones cambian y no hay necesidad de ahorrar agua o, incluso, conviene eliminarla, deja de secretarse ADH desde la neurohipófisis, baja su concentración sanguínea y, como consecuencia de ello, las aquaporinas son retiradas de la membrana apical de las células del epitelio del tubo colector. Disminuye así su permeabilidad al agua (llega a hacerse virtualmente impermeable) y deja de reabsorberse agua desde el interior del tubo hacia los espacios intersticiales. El volumen de orina es muy alto y su concentración osmótica, muy baja. De hecho, en seres humanos esa concentración puede ser tan baja como 50 mOsm, o sea, seis veces más baja que la plasmática, y el volumen de orina producido elevarse hasta representar un 15% del plasma filtrado en el glomérulo. Merece la pena reparar en el hecho de que una concentración osmótica de la orina tan baja no es solo el resultado de la supresión de la reabsorción de agua desde el tubo colector, sino que es necesario que se produzca una importante reabsorción de NaCl que no vaya acompañada de la correspondiente reabsorción de agua.
La ADH no es la única hormona implicada en la regulación de la función renal en mamíferos. La aldosterona y el péptido natriurético auricular cumplen también un importante papel. Antes de exponer brevemente en qué consiste, conviene advertir que tanto la aldosterona como, en general, las hormonas natriuréticas, o hormonas de similares naturaleza y efectos, se hallan en muchos otros grupos, además de mamíferos, pero su papel se entiende mejor en este contexto.
La aldosterona promueve la recuperación de Na+ desde la orina primaria y también la secreción de K+. Por ello, su efecto global más obvio es regular el contenido de esos iones en los fluidos corporales. Sin embargo, de forma indirecta también cumple un papel determinante en la regulación del volumen de los fluidos extracelulares, plasma sanguíneo incluido. La razón es que la concentración osmótica y de sales, como sabemos, se halla estrechamente controlada en los mamíferos y, en general, en el resto de los vertebrados. Por ello, el volumen de agua extracelular es muy dependiente de la cantidad de NaCl que hay en esos fluidos (no así en los intracelulares, cuyo catión principal es el K+), ya que Na+ y Cl– son los principales iones extracelulares. Así pues, si una hormona, como la aldosterona, promueve la reabsorción de Na+, también promueve la de Cl– y, por supuesto, la de agua. En otras palabras: cuanto más Na+ se reabsorbe en el riñón, también se reabsorbe más agua, por lo que su efecto neto es antidiurético.
La aldosterona es una hormona esteroidea, un mineralocorticoide producido por la corteza adrenal. Su secreción está controlada por otro sistema hormonal, el sistema renina-angiotensina que, a su vez, se encuentra parcialmente controlado por receptores de presión sanguínea y de volumen sanguíneo. Cuando la presión de la sangre baja y en virtud de varios mecanismos que actúan simultáneamente, las células yuxtaglomerulares (células especializadas del endotelio de la arteriola aferente) liberan renina. La acción (enzimática) de la renina sobre una molécula precursora de origen hepático (el angiotensinógeno) acaba dando lugar a que se produzca una sustancia denominada angiotensina II. Esta ejerce varios efectos: estimula la constricción (estrechamiento) de arteriolas sistémicas; promueve la sed; estimula la secreción de ADH y estimula la secreción de aldosterona. Todas las actuaciones provocadas por la angiotensina II causan la recuperación de agua y el reestablecimiento de la presión sanguínea y el volumen de líquidos extracelulares adecuados. Cuando la acción de la renina ha surtido sus efectos, ciertas sustancias paracrinas producidas por las células de la macula densa2 provocan que las células yuxtaglomerulares dejen de liberar renina.
La aldosterona actúa penetrando en sus células diana (a los efectos de lo que nos interesa aquí se trata de las células del epitelio del túbulo distal), llega al núcleo e inicia la transcripción de ADN para producir nuevas ATPasas de Na+-K+ y de canales de Na+ y de K+ para su inserción en la membrana celular. De esta forma se eleva la reabsorción tubular de Na+ y como consecuencia, la recuperación de agua y la restauración del volumen sanguíneo. Este mecanismo actúa, de hecho, en respuesta a situaciones –como las hemorragias- en las que se produce una importante pérdida de líquido sin que ello vaya asociado a una elevación de la concentración osmótica sanguínea.
Hay gran diversidad de péptidos natriuréticos en el dominio animal, y el mejor conocido es el péptido natriurético auricular (ANP, por sus siglas en inglés) de los mamíferos. El ANP se produce en determinadas zonas del encéfalo y en el corazón (de ahí su nombre “auricular”) y sus efectos son en gran parte opuestos a los de la aldosterona. Inhibe, de hecho, la liberación de aldosterona y promueve directamente la secreción de Na+, elevando la producción de orina y la concentración de Na+ en esta. La secreción de ANP es estimulada por el aumento del volumen de líquidos extracelulares, lo que es detectado a partir del estiramiento de las paredes de la aurícula en el corazón.
En definitiva, la función renal está sometida a un complejo sistema de regulación principalmente endocrino. Aquí hemos visto los tres sistemas principales, cada uno con sus especificidades. Gracias a ese sistema de efectos múltiples, los mamíferos, y demás vertebrados, son capaces de mantener estrechamente controlados tanto el volumen de los líquidos corporales, como sus concentraciones osmóticas e iónicas.
Notas:
1El mecanismo es muy similar al que vimos para la mayor parte de vertebrados, aunque en los otros grupos de vertebrados la ADH actúa en el túbulo distal y la molécula de efectos antidiuréticos en los demás vertebrados era algo diferente: arginina vasotocina (AVT).
2Grupo de células especializadas que se disponen en el punto en que el túbulo distal y la arteriola aferente se encuentran en posición adyacente.
Sobre el autor: Juan Ignacio Pérez (@Uhandrea) es catedrático de Fisiología y coordinador de la Cátedra de Cultura Científica de la UPV/EHU
Toman agua por la piel — Cuaderno de Cultura Científica
[…] los tubos colectores de las nefronas aumentando, precisamente, su permeabilidad al agua. Como vimos aquí, esos sistemas hormonales promueven el ahorro y la recuperación de agua, esto es, lo mismo que […]