Estructura y motilidad del intestino delgado

Estructura
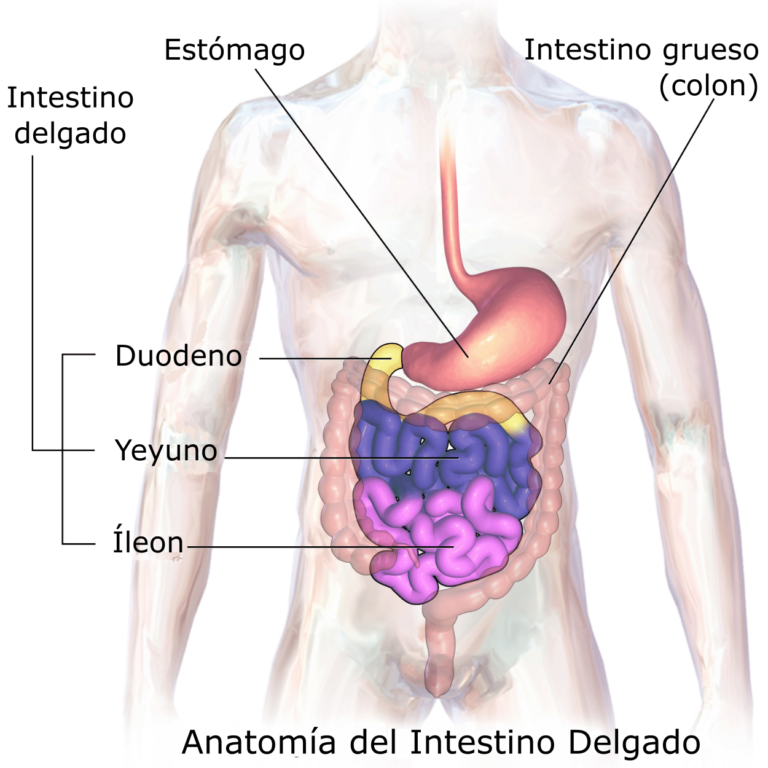
El intestino delgado es un tubo que se extiende desde el estómago hasta el intestino grueso. Se encuentra alojado en la cavidad abdominal, y recibe secreciones del páncreas y del sistema biliar. En la mayor parte de los vertebrados el intestino delgado es el principal órgano de digestión y absorción del alimento. En los terrestres se subdivide en tres áreas: duodeno, yeyuno e íleon.
Una sección transversal del tubo digestivo a la altura del intestino delgado revela la existencia de cuatro capas principales: mucosa, submucosa, muscular (muscularis externa) y serosa.
La mucosa se subdivide en tres láminas. La primera es el epitelio, denominado también membrana mucosa1, una lámina epitelial interior que contiene células exocrinas (secretan jugos digestivos), endocrinas (secretan hormonas) y epiteliales (especializadas en la absorción de nutrientes). La segunda es la lámina propia, una capa un tanto difusa de tejido conjuntivo en el que se insertan las células epiteliales de la membrana mucosa. La lámina propia se encuentra atravesada por finos vasos sanguíneos, conductos linfáticos y fibras nerviosas. Alberga además el tejido linfoide asociado al intestino, que es la barrera de defensa inmunitaria frente a los patógenos del intestino. La mucosa muscular es una fina capa de músculo liso que se encuentra entre la lámina propia y la submucosa.
La submucosa es una gruesa capa de tejido conjuntivo, al que debe la pared del intestino delgado su elasticidad. Contiene vasos sanguíneos y linfáticos cuyas ramificaciones se proyectan hacia la mucosa y hacia la capa muscular más externa. La submucosa también alberga una red nerviosa, llamada plexo submucoso o plexo de Meissner.
Por el exterior de la submucosa se encuentra la capa muscular. En la mayor parte del intestino delgado esta capa tiene dos subcapas, una interna, circular, y otra externa, longitudinal. La contracción de la subcapa interna provoca la constricción del tubo allí donde se produce, mientras que la contracción de la exterior provoca el acortamiento del tubo. La contracción combinada y coordinada de ambas subcapas es lo que produce la mezcla de los contenidos intestinales y su propulsión a lo largo del tubo. Entre ambas subcapas se encuentra otra red nerviosa, el plexo mientérico que junto al plexo submucoso ayuda a regular la actividad intestinal local.
La serosa es la capa de tejido conjuntivo que cubre el tubo digestivo. Secreta un fluido seroso de efecto lubricante que previene la fricción entre el aparato digestivo y los órganos adyacentes. No hay discontinuidad entre la serosa y el mesenterio que ancla el tubo digestivo a la pared de la cavidad abdominal.

La estructura del intestino delgado es tal que da lugar a un área superficial de gran extensión, muy superior a la que tendría un simple tubo de la misma longitud y diámetro luminal. La existencia en la mucosa de pliegues, vellosidades y microvellosidades es lo que permite que la superficie interior sea muy superior a lo que cabría esperar de un tubo sin esas particularidades, y gracias a ello el área disponible para la absorción alcanza una gran magnitud.
La superficie interior de la mucosa se dispone en pliegues circulares que multiplican por tres la superficie interna del tubo. De los pliegues salen proyecciones similares a dedos microscópicos; son los villi (villus en singular) o vellosidades, y dan una apariencia aterciopelada a la superficie interna del intestino. Multiplican por diez el área superficial. La superficie de cada villus se halla cubierta por células epiteliales, de las que salen las microvellosidades (o microvilli); estas forman lo que se denomina borde en cepillo (brush border). Cada célula epitelial puede contener en su parte apical entre 3000 y 6000 microvillosidades, y permiten multiplicar por veinte el área superficial de las células. El borde en cepillo alberga enzimas que participan en la digestión y la absorción simultánea de sus productos. En total, la superficie efectiva para la absorción es seiscientas veces mayor de lo que sería en un tubo cuya pared interna fuese lisa.
Las uniones estrechas (o zonulae occludentes) entre las células epiteliales de los villi mantienen herméticamente aislados la luz intestinal y el medio intersticial. La absorción se produce gracias al concurso de transportadores específicos de cada nutriente o electrolito en el borde en cepillo, y a la acción de las enzimas allí insertas que completan la digestión de carbohidratos y proteínas.
Por otro lado, cada villus recibe una arteriola que se ramifica en una red de capilares en su interior. Además, el centro de cada vellosidad está ocupada por un vaso linfático ciego que denominado vaso quilífero. La absorción consiste en la transferencia de los nutrientes digeridos a los capilares y al vaso linfático terminal, para lo que han de atravesar las células epiteliales de la mucosa, difundir a través del fluido intersticial que baña el tejido conjuntivo del núcleo de las vellosidades y atravesar el endotelio de algún capilar o del vaso quilífero.
Motilidad
Al llegar el quimo al duodeno, las contracciones de la musculatura lisa provocan su mezcla con las secreciones procedentes de páncreas e hígado, y lo impulsan a lo largo del tubo. La forma primaria de motilidad es la segmentación, proceso que consiste en contracciones anulares de la muscula lisa circular a lo largo del intestino delgado. Las contracciones obliteran el tracto intestinal, de manera que entre cada dos zonas contraídas, las relajadas albergan porciones discretas de quimo. Esas contracciones no se desplazan de la forma en que lo hacen las peristálticas, sino que se alternan con momentos de relajación. Contracciones y relajaciones se suceden en cada zona, lo que provoca que el quimo que se hallaba entre dos zonas contraídas se divida en dos partes y se mezcle con el de las zonas adyacentes. La reiteración de esa secuencia de contracciones y relajaciones sucesivas da lugar a una mezcla completa del contenido intestinal. Además, de esa forma se expone todo ese contenido a la superficie interna de la mucosa, lo que permite la absorción homogénea de todos los nutrientes.
Las contracciones de la segmentación empiezan debido a la acción de las células marcapasos del intestino delgado, células que generan un ritmo eléctrico básico similar al del estómago. Si el potencial eléctrico que produce esas células supera un determinado umbral, provoca la contracción de la capa de musculatura lisa circular a la frecuencia propia de las células marcapasos.
La intensidad de la respuesta de la musculatura no es constante, por lo que la intensidad de las contracciones de segmentación depende del grado de distensión de la pared intestinal, de la acción de la gastrina y de la acción nerviosa extrínseca. Entre comidas, la segmentación es mínima o, sencillamente, no se produce. Pero cuando llega la comida al tracto digestivo se producen fuertes contracciones de segmentación. Las contracciones duodenales empiezan en respuesta a la distensión producida por la presencia de quimo procedente del estómago. Pero las que se producen más adelante, en el íleon, son estimuladas por la gastrina, que se libera en respuesta a la presencia de quimo en el estómago. Además, la estimulación parasimpática (y por lo tanto, extrínseca), refuerza la segmentación, mientras que la simpática provoca el efecto contrario.
La progresión del quimo a lo largo del intestino delgado no se produce mediante peristaltismo, como ya se ha señalado. El quimo avanza porque la frecuencia de la segmentación se reduce a lo largo de su recorrido. De lo contrario no ocurriría, dado que las contracciones provocan el desplazamiento del quimo tanto hacia delante como hacia atrás. Las células marcapasos del duodeno se despolarizan espontáneamente con una frecuencia mayor que las más alejadas, siendo las del íleon terminal las que lo hacen a menor frecuencia. Las segmentaciones del intestino delgado humano pasan así de ser 12 por minuto en el duodeno a solo 9 por minuto en el íleon terminal. Esa diferencia hace que se desplace algo más quimo hacia delante que hacia atrás, dando lugar a un lento avance. De ese modo se va produciendo la mezcla a la vez que los nutrientes son absorbidos, habiendo tiempo suficiente para ello. Normalmente, el quimo tarda entre 3 y 5 horas en recorrer el intestino delgado humano.
Cuando ya se ha absorbido la mayor parte del alimento la segmentación cesa y es sustituida por lo que se denomina complejos mioeléctricos migratorios (o complejo motor migrante, CMM), también denominados “amo de casa intestinal”. Consiste este complejo en ondas de actividad eléctrica que se desplazan a lo largo del intestino delgado entre comidas. Las ondas eléctricas generan contracciones peristálticas. Y de esa forma, los restos de alimento no digerido, junto con bacterias, sustancias difíciles de digerir y restos de mucosa son transportadas hasta la válvula ileocecal y el interior del colon para su expulsión final como restos fecales. Un complejo motor migrante se desarrolla a lo largo de unas dos horas, y una vez finalizado vuelve a producirse hasta que vuelve a llegar quimo al estómago.
La válvula ileocecal permanece abierta mientras los contenidos del intestino delgado son impulsados a su través hacia el intestino grueso. Pero se cierra ante la mínima señal que amenace con retrotraer los restos de la digestión al intestino delgado. Por otro lado, la musculatura lisa del último tramo del íleon se encuentra engrosada y, por lo tanto, en condiciones, al contraerse, de formar un esfínter que se encuentra bajo control hormonal y nervioso. Normalmente ese esfínter se encuentra prácticamente cerrado, y se cierra con más intensidad si hay alguna presión en el lado del intestino grueso; por el contrario, si el lado del íleon se encuentra distendida, entonces ese esfínter se mantiene abierto. Mediante esos dos mecanismos (válvula ileocecal y musculatura lisa engrosada) permiten controlar el posible retroceso de los restos de alimento no digerido y, lo que es más importante, la invasión de patógenos procedentes del intestino grueso.
Nota:
1 Aunque esa denominación se presta a confusión porque a la mucosa también se la suele denominar así.
Sobre el autor: Juan Ignacio Pérez (@Uhandrea) es catedrático de Fisiología y coordinador de la Cátedra de Cultura Científica de la UPV/EHU
El intestino grueso — Cuaderno de Cultura Científica
[…] de las células musculares lisas del colon, y son similares a las contracciones que provocan la segmentación en el intestino delgado, solo que se producen con una frecuencia muy inferior. Se producen entre 9 y 12 contracciones de […]
El intestino grueso – Fluceando
[…] de las células musculares lisas del colon, y son similares a las contracciones que provocan la segmentación en el intestino delgado, solo que se producen con una frecuencia muy inferior. Se producen entre 9 y 12 contracciones de […]
UNIDAD III. BALANCE ENERGÉTICO | 5FM1 FISIOLOGÍ HUMANA
[…] las células musculares lisas del colon, y son similares a las contracciones que provocan la segmentación en el intestino delgado, solo que se producen con una frecuencia muy inferior. Se producen entre 9 y 12 contracciones de […]