Digestión estomacal (y II): regulación
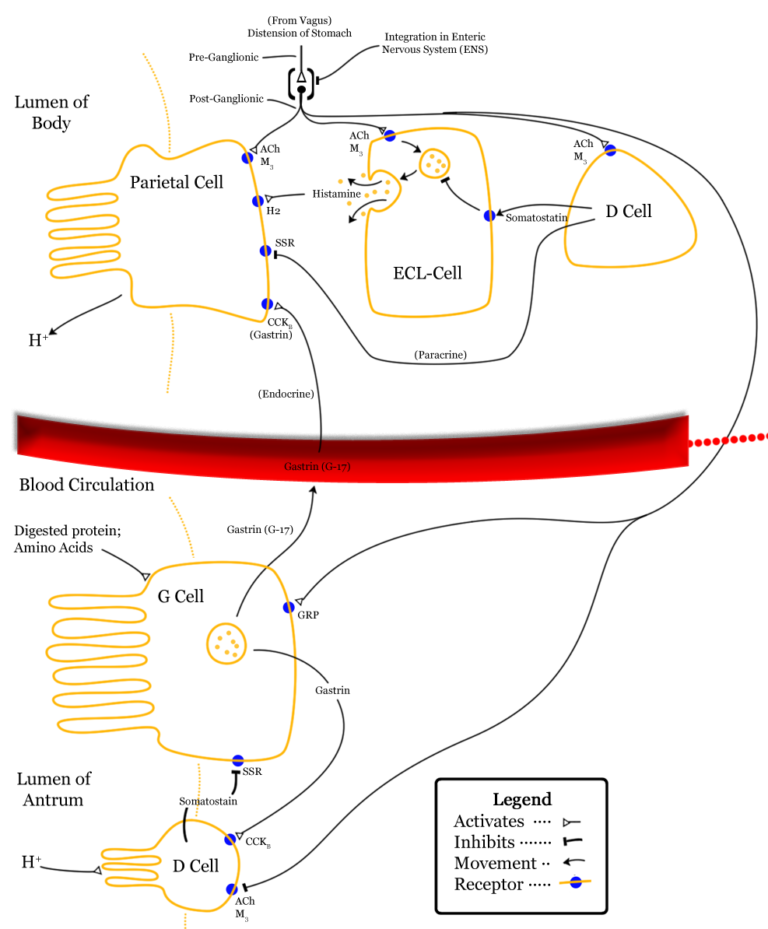
Además de las células propias de las glándulas de la mucosa oxíntica que se presentaron en la anotación anterior, en la mucosa gástrica hay otras que no liberan productos exocrinos, sino factores endocrinos (hormonas) o paracrinos con función reguladora. Son las siguientes:
(1) Las células enterocromafines se encuentran entre las células parietales y las células principales en las glándulas gástricas de la mucosa oxíntica. Secretan histamina.
(2) Las células G se encuentran en las glándulas del área de glándulas pilóricas. Son células endocrinas, que liberan la hormona gastrointestinal gastrina a la sangre.
(3) Las células D (o delta) se encuentran en número escaso en las glándulas de la zona pilórica pero son muy abundantes en el duodeno. Secretan somatostatina.

La secreción de los jugos gástricos que vimos en la anotación anterior se produce mediante la intermediación de cuatro mensajeros químicos, para los que las células parietales cuentan con sus correspondientes receptores. Acetilcolina, gastrina e histamina ejercen efectos estimulantes al promover la inserción de las unidades de ATPasa de H+/K+ -que se encuentran almacenadas en vesículas intracelulares- en la membrana de las células parietales, elevando así la secreción de HCl a la luz gástrica. La somatostatina, sin embargo, tiene efectos inhibitorios. La gastrina y la acetilcolina estimulan también la secreción de pepsinógeno por las células principales. Veamos estos mensajeros con algún detalle.
La acetilcolina es un conocido neurotransmisor. Se libera en el plexo nervioso intrínseco en respuesta a la estimulación por el nervio vago, y estimula las células parietales, principales, células G y las enterocromafines.
Las células G secretan gastrina al torrente sanguíneo, en respuesta a la presencia de proteínas en el interior del estómago y por efecto de la acetilcolina. Cuando vuelve al cuerpo y al fundus del estómago, las células parietales y principales elevan la secreción de jugos gástricos. Además, promueve la liberación de histamina por parte de las células enterocromafines.
La histamina es un agente paracrino liberado por las células enterocromafines en respuesta a la gastrina y la acetilcolina. Ejerce efectos locales (por eso es paracrina) en las células parietales de su entorno, potenciando la acción, precisamente, de las sustancias que han estimulado su liberación.
La somatostatina es liberada por las células delta (o células D) en respuesta a la acidez y su efecto es inhibitorio dando lugar a una retroalimentación negativa. Actúa como agente paracrino también, sobre las células de su entorno, inhibiendo la secreción de células parietales, células G y células enterocromafines.
La regulación de la digestión estomacal se produce mediante un conjunto de actuaciones complementarias. Son varios los estímulos capaces de provocar un aumento de la secreción de jugos gástricos. Unos son anteriores a la llegada del alimento a estómago, otros actúan cuando el alimento ya se encuentra en su interior, y otros proceden del duodeno una vez el quimo ha abandonado el estómago. Por esa razón la secreción gástrica sigue una secuencia en tres fases:
Fase cefálica. Antes de que el estómago reciba el alimento, determinados estímulos sensoriales (visuales, olfativos o el acto de masticar) anticipan su llegada. En ese caso la secreción gástrica se eleva por la acción del nervio vago en respuesta a los citados estímulos, acción que surte dos efectos: (1) promueve el aumento en la secreción de acetilcolina por los plexos nerviosos intrínsecos, lo que, a su vez, eleva la secreción de HCl y de pepsinógeno; (2) estimula la secreción de gastrina por las células G, lo que a su vez promueve la secreción de HCl y pepsinógeno; además, la secreción de clorhídrico también se eleva debido a que la gastrina induce la liberación de histamina.
Fase gástrica. Esta comienza al llegar el alimento al estómago. Los estímulos que actúan a este nivel (proteínas y péptidos, por un lado, y distensión estomacal por el otro) elevan la secreción gástrica debido a la acción superpuesta de diferentes vías. La acción más potente se debe a la presencia de proteínas en el estómago, que estimulan los quimiorreceptores que activan los plexos nerviosos intrínsecos, con la consiguiente estimulación por estos (mediante la acetilcolina) de la células secretoras. La presencia de proteínas en el estómago también actúa a través de la vía vagal, reforzando la fase cefálica. La consecuencia de estas acciones es la elevación de la secreción de ácido clorhídrico y de pepsinógeno, acentuándose así la digestión ya iniciada.
Fase intestinal. Hay factores intestinales que inciden en la secreción de jugos gástricos, aunque en este caso pueden ser estimuladores o inhibidores. La presencia en el duodeno de los productos de la digestión parcial de las proteínas estimula la liberación de gastrina intestinal, que es transportada al estómago por la sangre, actuando de la misma forma que la gastrina estomacal. Ahora bien, cuando el quimo llega las zonas iniciales del intestino delgado la secreción gástrica se reduce debido a tres factores:
(1) Por un lado, al vaciarse el estómago, disminuye la presencia de proteínas en su interior, desapareciendo de esa forma el principal factor que había provocado la elevación de la secreción de jugos.
(2) Conforme se vacía el estómago, su pH se reduce mucho, pues el HCl ya no es neutralizado por las sustancias del quimo; esa reducción estimula la liberación de somatostatina que, como hemos visto, ejerce efecto inhibidor sobre la secreción gástrica.
(3) Los mismos estímulos que inhiben la motilidad estomacal, inhiben también la secreción gástrica. El reflejo enterogástrico y las enterogastronas suprimen la actividad de las células secretoras gástricas, a la vez que reducen la excitabilidad de las células musculares lisas.
Llegados a este punto, la digestión estomacal ha terminado y sus productos han ido pasando del antro estomacal al duodeno e intestino delgado, donde se completará la digestión de los péptidos, lípidos y carbohidratos que no han sido hidrolizados en las fases anteriores.
Sobre el autor: Juan Ignacio Pérez (@Uhandrea) es catedrático de Fisiología y coordinador de la Cátedra de Cultura Científica de la UPV/EHU
Hector04
excelente artículo, una consulta y que papel juegan las neuronas esparcidas por el sistema digestivo en este proceso
Juan Ignacio Pérez Iglesias
Sobre el papel de las neuronas (no diría que esparcidas, ya que forman parte de un sistema organizado en plexos) algo se ha dicho, dentro de esta misma serie, aquí: https://culturacientifica.com/2018/10/22/actividades-digestivas-y-su-regulacion/ y aquí: https://culturacientifica.com/2018/12/31/la-motilidad-estomacal/
Y sobre el sistema entérico en su conjunto, Pablo Barrecheguren escribió también esto: https://culturacientifica.com/2016/12/09/intestino-no-segundo-cerebro/