Absorción del alimento
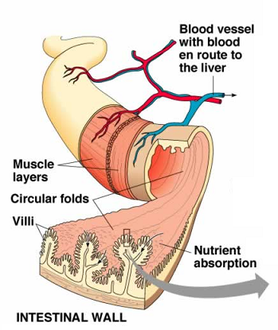
La mucosa del intestino delgado de los mamíferos contiene glándulas exocrinas cuyo producto de secreción es una solución de moco y sal denominada succus entericus o jugo intestinal. La función de esa secreción es aportar un agente protector y lubricante de la pared intestinal (el moco) y agua, ya que la digestión requiere que los nutrientes se encuentren en disolución.
Las enzimas que realizan la digestión en la luz del intestino provienen del páncreas. Son la lipasa, tripsina, quimotripsina, carboxipeptidasa y amilasa. Los productos de esa digestión o bien son absorbidos directamente o son digeridos por enzimas del borde en cepillo de las células epiteliales. Las enzimas en cuestión son la enteroquinasa (que activa el tripsinógeno procedente del páncreas), las disacaridasas que finalizan la digestión de los carbohidratos (en mamíferos estas enzimas son maltasa, sacarasa, lactasa y trehalasa) y las aminopeptidasas, que hacen lo propio con la digestión de las proteínas. En las aves hay alguna diferencia con respecto a los mamíferos: carecen de lactasa, como es lógico, y muchos pájaros (no así los colibríes) no tienen sacarasa.
En principio, el intestino tiende a absorber todos los nutrientes (tras su digestión), electrolitos1, vitaminas y agua que han sido ingeridos. Aunque, como veremos en otra ocasión, es un órgano muy flexible, por lo que la capacidad para digerir y absorber unos nutrientes u otros se puede modificar en función de su presencia y proporción en la dieta habitual.
El agua se absorbe de forma pasiva impulsada por la absorción de Na+. Este, por su parte, puede pasar a favor de gradiente electroquímico –y, por lo tanto, de forma pasiva- desde la luz del tubo al fluido intersticial que hay entre los villi. Ese movimiento se produce a través de los espacios intercelulares; del líquido intersticial pasa a los capilares sanguíneos.
Pero también hay movimiento de sodio a través de las células epiteliales, y ese movimiento sí requiere gasto de energía. Puede entrar pasivamente en la célula a través de canales o en compañía de otro ión gracias al concurso de un transportador (simporter Na+/Cl–, antiporter Na+/H+ o simporter Na+-glucosa (o aminoácido)). Y sale de la célula epitelial, a través de su membrana basolateral hasta el líquido intersticial de los espacios intercelulares gracias a la acción de la bomba Na+/K+. La absorción de Cl–, H2O, glucosa y aminoácidos está vinculada a la absorción activa de sodio. El Cl– pasa a favor del gradiente eléctrico creado por el Na+, y el agua, por el flujo osmótico provocado por la alta concentración de NaCl en los espacios intersticiales.
La mayor parte de los carbohidratos en condiciones de ser absorbidos por las células epiteliales se encuentran, como ya se ha dicho, en forma de disacáridos. Las disacaridasas presentes en el borde en cepillo reducen esos azúcares a sus monómeros constituyentes: glucosa, galactosa y fructosa, dependiendo del disacárido original.
Glucosa y galactosa son absorbidas mediante transporte secundario activo, en el que intervienen simporters como el cotransportador sodio-glucosa (SLGT), que transfieren ambas sustancias, glucosa y sodio desde la luz del intestino al interior celular. La energía que se gasta en este proceso es la que gasta la bomba Na+/K+ de la membrana basolateral que, como antes hemos señalado, es la responsable de crear el gradiente de Na+ necesario para hacer posible su transferencia y, con él, el del azúcar. De esa forma, la concentración de glucosa (y en su caso la de galactosa) se eleva dentro de la célula, por lo que puede abandonarla fácilmente a través de la membrana basal a favor de gradiente de concentración, mediante difusión facilitada por el transportador de glucosa GLUT-2, presente en esa membrana. Del fluido intersticial pasa al capilar del villus en el que se encuentra la célula epitelial. Además de ese transporte, parece ser que la glucosa también puede pasar desde la luz intestinal hasta el líquido intersticial a través de los espacios intercelulares del epitelio (como ocurría con el sodio).
La fructosa se absorbe a través de difusión facilitada. Entra en la célula epitelial gracias al transportador GLUT-5 y sale al espacio intersticial mediante el concurso del transportador GLUT-2.
Las proteínas, por su parte, se van fragmentando, primero en el estómago y luego en la luz intestinal. Como resultado de las sucesivas hidrólisis anteriores, finalmente quedan aminoácidos y pequeños péptidos para su digestión final y absorción epitelial. Los aminoácidos se absorben de forma similar a la glucosa y galactosa.
Los pequeños péptidos se introducen en la célula epitelial mediante un transportador dependiente de Na+. El proceso se denomina transporte terciario activo. El simporter que interviene transfiere a la vez el péptido y un protón desde la luz intestinal al interior de la célula. El H+ se mueve a favor de gradiente de concentración, mientras que el péptido lo hace en contra. El gradiente de protones se genera mediante el concurso de un antiporter de Na+/H+, que introduce sodio a favor de gradiente de concentración a la vez que saca H+ en contra. Por su parte, el gradiente de concentración de Na+ que impulsa el antiporter Na+/H+ se genera gracias a la actividad de la bomba Na+/K+ en la membrana basolateral.
Todo ese proceso se puede observar desde el ángulo contrario. La bomba Na+/K+ extrae Na+ de la célula hacia el líquido intersticial. Ese transporte es necesariamente activo, porque el sodio está muy concentrado en el medio interno (el líquido intersticial o extracelular es medio interno), mientras que el potasio lo está en el medio intracelular. La bomba, por lo tanto, reduce de esa forma la concentración intracelular de Na+, de manera que facilita su transferencia desde la luz intestinal. Dado que parte de ese sodio entra acoplado a la salida de protones y puesto que esa salida está, a su vez, asociada a la entrada de oligopéptidos a la célula, la incorporación de estos acaba siendo el resultado de una cadena de movimientos, activos y pasivos, en la que intervienen varios iones (H+, Na+ y K+).
Los oligopéptidos son hidrolizados por la acción de las aminopeptidasas del borde en cepillo de la membrana celular o en el citoplasma gracias a la acción de peptidasas intracelulares. Los aminoácidos resultantes de estos procesos (transporte o digestión intracelular) salen de la célula epitelial mediante transportadores pasivos (no utilizan energía) y entran en los capilares sanguíneos por difusión.
Además de las proteínas provistas por el alimento, en el intestino delgado se digieren y absorben proteínas de origen endógeno, como enzimas digestivas, proteínas celulares que han sido expulsadas de las células de la pared a la luz intestinal durante la renovación de la mucosa, y proteínas plasmáticas que se filtran desde el plasma accidentalmente. Entre 20 y 40 g de estas proteínas son digeridas diariamente.
Las grasas experimentan un proceso de digestión y absorción muy diferente de los anteriores. Como vimos, debido a la acción de las sales biliares y los movimientos intestinales las gotas lipídicas de tamaño grande se convierten, primero en gotas pequeñas, y luego, debido a la acción de la lipasa (con el concurso de la colipasa), en micelas, estructuras lipídicas esféricas de entre 3 y 10 nm de diámetro. Esas micelas son la forma en que los monoglicéridos y ácidos grasos libres acceden a la membrana de las células epiteliales para, una vez allí, penetrar a su través. Los componentes que configuran la estructura micelar son reutilizados para englobar nuevos fosfolípidos y monoglicéridos, y llevarlos hasta la membrana para su absorción.
Tras entrar en las células, los constituyentes de los lípidos son utilizados para sintetizar triglicéridos, que son empaquetados en agregados provistos de una capa de lipoproteínas gracias a la cual se hacen solubles en agua. Las lipoproteínas de mayor tamaño se denominan quilomicrones, y salen mediante exocitosis de la célula epitelial hacia el líquido intersticial, y de ahí, en mamíferos, al vaso quilífero (al que nos referimos aquí). En las aves los quilomicrones pasan a los capilares sanguíneos de los villi.
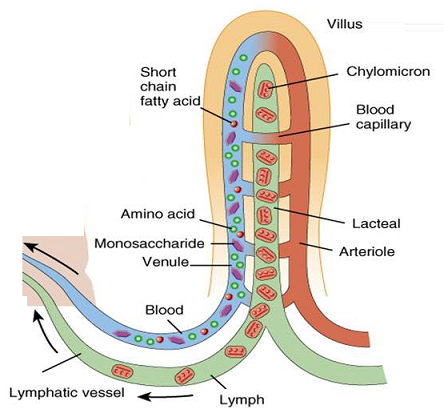
Los productos de la digestión de carbohidratos y proteínas pasan al hígado porque, como vimos, las vénulas desembocan en la vena porta hepática. La mayor parte de esos nutrientes se procesan metabólicamente allí. Además, ese órgano también se ocupa de tareas de desintoxicación de sustancias dañinas. Las grasas tienen un destino diferente. Al acceder al vaso quilífero, acaban desembocando en el sistema linfático y, de este, pasan a la circulación general a través del conducto torácico, un gran vaso linfático que desemboca en el sistema venoso. Ese dispositivo hace que una parte importante de los lípidos absorbidos sean incorporados directamente a los adipocitos, aunque otra parte acaba también siendo procesada por el hígado. De este modo se evita que este órgano reciba más grasa de la que puede procesar.
El hígado, no obstante, juega una función importante en el transporte de lípidos pues, como vimos, sintetiza diferentes lipoproteínas plasmáticas y las vierte al sistema circulatorio para su uso. Las lipoproteínas de alta densidad (HDL y LDL) transportan principalmente colesterol y también fosfolípidos para la fabricación de membranas celulares. Las lipoproteínas de muy baja densidad (VLDL) transportan principalmente triglicéridos que son almacenados en adipocitos como sustancias de reserva.
Hemos visto antes que la absorción de sodio por las células de la mucosa intestinal va acompañada de absorción de agua. Es, de hecho, una actividad muy importante. Un intestino delgado humano viene absorber del orden de 9 l de agua al día, aunque el agua que se ingiere en el alimento son unos 1.250 ml y otros 1.250 ml se ingieren bebiendo. El resto son fluidos procedentes del propio organismo: 1.500 ml de saliva, 2.000 ml de jugos gástricos, 1.500 ml de jugo pancreático, 500 ml de bilis y 1.500 de jugo intestinal. La suma de esas cantidades arroja un volumen de 9.500 ml. Los 500 ml restantes pasan al intestino grueso, donde se absorberán 350 ml diarios. El resto, 150 ml, se evacúa con las heces.
Por otra parte, el HCl secretado por el estómago es neutralizado en el intestino delgado por el NaHCO3 secretado por el páncreas. El H2CO3 resultante se disocia en CO2 y H2O y estos productos, junto con el NaCl también resultante de la neutralización del clorhídrico procedente del estómago son absorbidos. El resultado final es que las actividades digestivas no provocan cambios en el pH del medio interno.
Nota:
1Con la salvedad de calcio y hierro, que son absorbidos en función de las necesidades.
Sobre el autor: Juan Ignacio Pérez (@Uhandrea) es catedrático de Fisiología y coordinador de la Cátedra de Cultura Científica de la UPV/EHU
Absorción del alimento | Hacia una Cultura Científica
[…] https://culturacientifica.com/2019/04/08/absorcion-del-alimento/ […]
Diferencia entre Alimentación y Nutrición | LasDiferencias.wiki
[…] La nutrición es la ingesta de alimentos y el uso de los nutrientes de los alimentos en el cuerpo. Luego de la ingesta alimentaria que viene a ser la primer etapa de la nutrición, el sistema desarrolla una serie de procesos posteriores en donde los alimentos son descompuestos para su absorción. […]