Los pulmones de reptiles y aves

Esta anotación debería quizás llevar el nombre “saurópsidos1” en el título, pero entonces pocos habrían sabido de qué se trata. Por eso he optado por referirme a reptiles y aves. Hay gran variedad de pulmones en los saurópsidos. Los más sencillos son sacos simples en los que prácticamente no hay compartimentos. De hecho, se podría decir de un pulmón así que es como un único gran alveolo. También hay pulmones más complejos, con numerosos septos o paredes que se proyectan desde la superficie pulmonar hacia la luz. Los septos delimitan unos cubículos, unidades espaciales denominadas edículas. El intercambio de gases tiene lugar, principalmente, en los septos, aunque no están tan vascularizados como los alveolos de mamíferos. En otros casos los pulmones son multicamerales; están constituidos por diferentes cámaras dispuestas en paralelo. En estos, un bronquio intrapulmonar principal se va ramificando en bronquios secundarios que conectan el bronquio principal con cada cámara o saco aéreo.
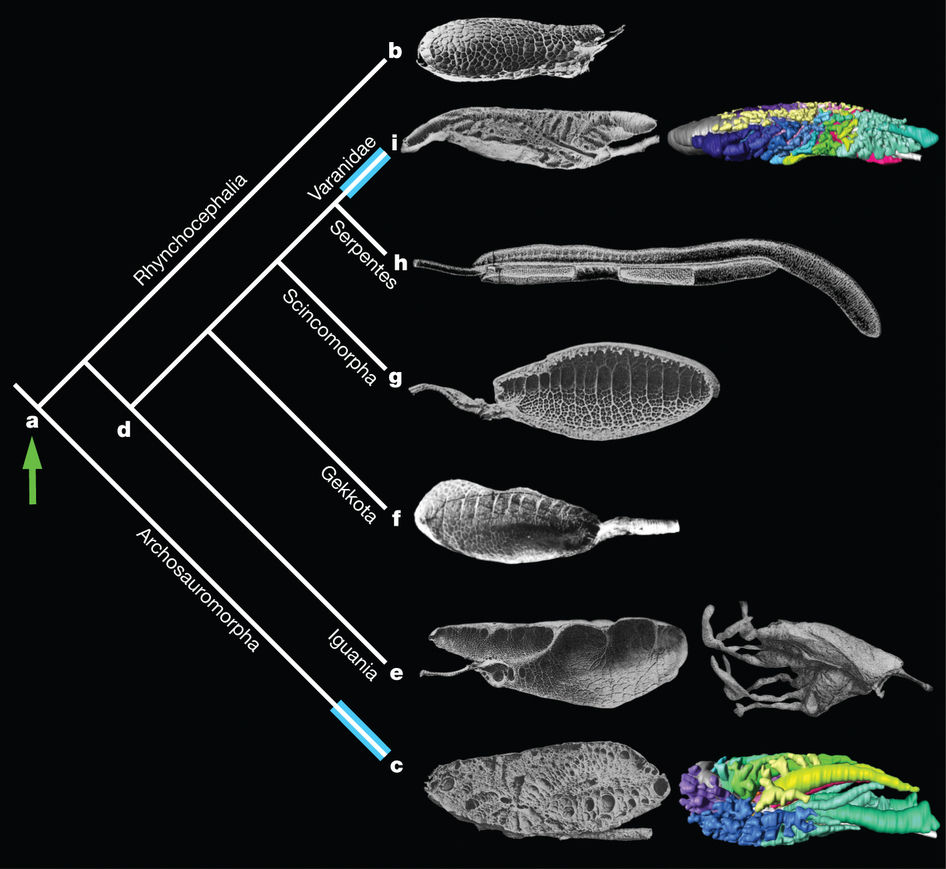
En la gran mayoría de reptiles estudiados, la ventilación es mareal: se produce por flujo y reflujo del aire respirado, como si fuese impulsado por un fuelle. Una parte importante de muchos pulmones de reptiles no se dedica al intercambio respiratorio, sino que su función principal es ayudar a ventilar las porciones bien vascularizadas. Los mecanismos que participan en la impulsión del aire que entra y sale de los pulmones de los reptiles son variados, pero en general corren a cargo de la musculatura del tronco; como, en ocasiones, esa musculatura también participa en los movimientos de desplazamiento, ello puede representar una limitación. Quizás por esa razón algunos, como los varanos y otros lagartos, también utilizan la boca como bomba de impulsión del aire, como hacen las ranas. Otros reptiles poseen un diafragma o protodiafragma, cuyos desplazamientos arrastran el hígado hacia atrás y hacia adelante, dando lugar a la expansión y contracción de los pulmones en función del espacio que deja aquel.
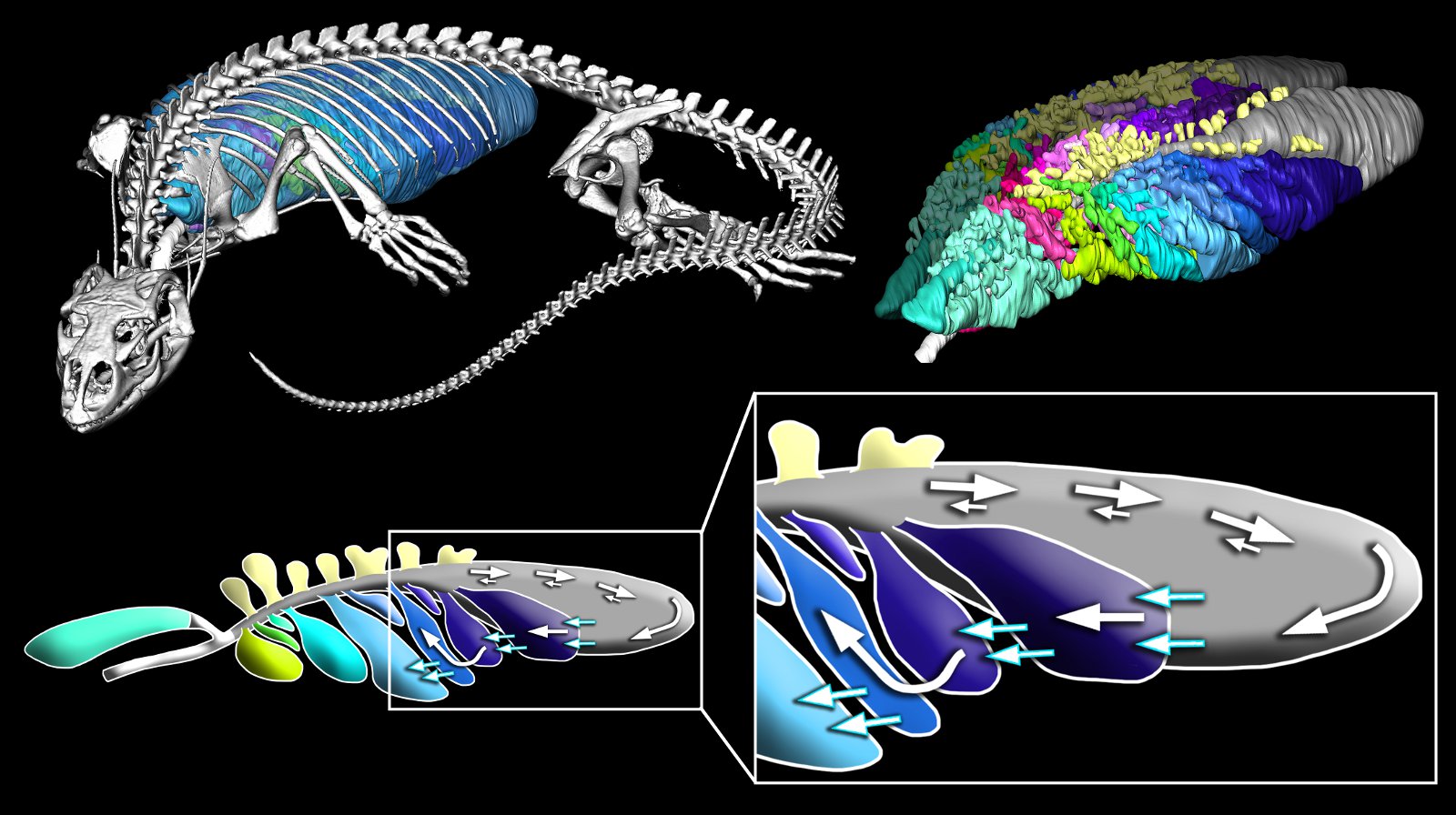
Algunos reptiles, como es sabido, tienen un modo de vida acuático. Entre estos, unas especies han retenido la condición de respiradores aéreos, como los cocodrilos, algunas tortugas y las serpientes de mar, a pesar de lo cual pueden hacer inmersiones prolongadas. Hay serpientes de agua dulce capaces permanecer sumergidas hasta media hora; la tortuga verde marina llega a estar bajo el agua 50 minutos; y las serpientes marinas aguantan sin respirar una hora. Estas tienen un pulmón muy largo, que se extiende a lo largo de casi todo su cuerpo, por lo que es capaz de almacenar mucho oxígeno; además, reducen de forma considerable su metabolismo durante la inmersión. Y hay especies que tienen la capacidad, al sumergirse, de recurrir a vías metabólicas anaerobias para obtener ATP.
Otros reptiles son capaces de incorporar oxígeno del medio acuático de forma directa, como algunas culebras y tortugas. Las serpientes lo hacen a través de su superficie corporal, mientras que las tortugas recurren en mayor medida a captar O2 a través de las cavidades bucal y faríngea, e incluso, a través de la cloaca, que ha sufrido modificaciones que facilitan el intercambio de gases respiratorios. Ninguna de esas superficies es demasiado extensa, de manera que el intercambio gaseoso a su través es muy limitado, pero parece ser suficiente para cubrir las necesidades metabólicas de esos animales. Al fin y al cabo, dado que se trata de animales ectotermos, su metabolismo es relativamente bajo.
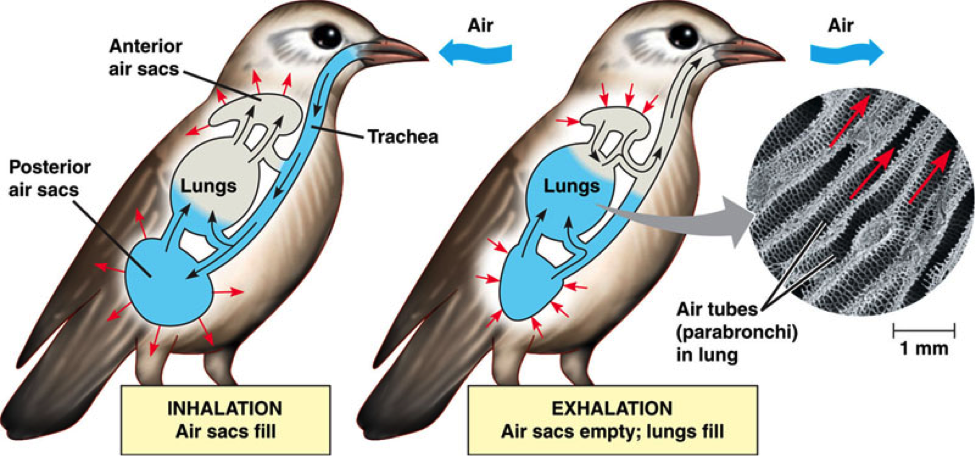
Los pulmones de las aves, aunque relativamente pequeños, son los más complejos. En cada pulmón, el bronquio principal se ramifica en varios bronquios secundarios, y estos están conectados entre sí por numerosos bronquios terciarios o parabronquios. Los parabronquios son cilíndricos y discurren en paralelo unos con otros. Los bronquios secundarios y los parabronquios forman una unidad integrada que permite que el aire circule a su través siempre en la misma dirección; el aire pasa de los llamados dorsobronquios a los ventrobronquios (denominaciones relativas a la posición que ocupan en el pulmón) a través de los parabronquios. La corriente inhalante entra por los dorsobronquios y la exhalante sale por los ventrobronquios. Al pasar por los parabronquios, el aire penetra en los capilares aéreos, un conjunto de canalículos interconectados que forman una tupida red y que aumentan de forma muy considerable la superficie para el intercambio gaseoso. Los capilares aéreos están a su vez íntimamente entrelazados con capilares sanguíneos en una disposición denominada de corriente cruzada (cross current), de manera que la sangre circula en una dirección que mantiene un ángulo de unos 90º, aproximadamente, con la dirección que sigue el aire respirado. Esa disposición permite una transferencia de O2 de altísima eficiencia.
Los pulmones de las aves están conectados con un conjunto de sacos aéreos, unos en disposición anterior y otros, posterior. Los sacos posteriores reciben el aire recién inhalado, de allí pasa a los pulmones y de estos, a los sacos anteriores, de donde es expulsado al exterior. Los sacos están dispuestos de manera que el aire inhalado no se mezcla con el que ya ha pasado por los parabronquios, por lo que todo el aire que llega al pulmón es aire fresco. El aire circula a través de ese sistema gracias, principalmente, a la contracción de los músculos de las costillas en asociación con el esternón.
Las pulmones de aves son, sin duda, los órganos respiratorios más eficaces de entre los metazoos. Ello se debe, como se ha señalado ya, a la circulación unidireccional de la corriente respiratoria y a la disposición en corriente cruzada de los parabronquios y los capilares sanguíneos. En esos rasgos las aves se diferencian de forma clara de la mayor parte del resto de saurópsidos. Sin embargo, es importante destacar que hay, al menos, tres especies de saurópsidos no aviares (un cocodrilo, un varano y una iguana) en cuyos pulmones hay flujo de aire unidireccional. Y es muy posible que otras especies también cuenten con esa ventaja, o que otras ya extinguidas también la tuviesen. Así pues, aunque el pulmón de las aves tiene características muy sofisticadas, todo hace indicar que hubo versiones previas, aunque quizás no fuesen tan eficaces.
Nota:
1 El clado Sauropsidae comprende todos los denominados reptiles, salvo los extinguidos “reptiles mamiferoides” (supuestos antecesores de los mamíferos) y las aves. La razón por la que quizás debería haber optado por ese nombre en el título es que los pulmones de las aves tienen seguramente antecedentes en otros saurópsidos.
Sobre el autor: Juan Ignacio Pérez (@Uhandrea) es catedrático de Fisiología y coordinador de la Cátedra de Cultura Científica de la UPV/EHU

Luna Villar Romero
No hay pingüinos nesesito saber de que tamaño son los pulmones de el pingüino Común gracias por leer este comentario Luna de 9 años. Y gracias
Una nueva (e inesperada) función de los sacos aéreos — Cuaderno de Cultura Científica
[…] respiratorio debe suministrar oxígeno en grandes cantidades. Esta es la razón por la que el sistema respiratorio de las aves es completamente diferente al de cualquier otro grupo de vertebrados, incluyendo a los mamíferos. […]