La regulación osmótica e iónica en los teleósteos marinos

La concentración osmótica del medio interno de los teleósteos marinos está muy por debajo de la concentración del medio en el que viven. El agua de mar tiene una osmolaridad de unos 1000 mOsm (miliosmolar) y la de los peces óseos se encuentra normalmente entre 300 y 400 mOsm, aunque en algunos casos puede ser algo mayor. La diferencia, como se puede apreciar, es muy importante. De hecho, son valores muy próximos a los de los teleósteos de agua dulce (entre 250 y 350 mOsm), proximidad que refleja el común origen dulceacuícola de todos ellos.
Los teleósteos marinos son, por lo tanto reguladores osmóticos, como lo son los de agua duce; lo que ocurre es que estos son reguladores hiposmóticos, puesto que mantienen la concentración interna por debajo de la exterior. Dada su condición hiposmótica con respecto al medio en el que se encuentran, han de hacer frente al riesgo de desecación que tal condición comporta. En estos animales el agua tiende a salir del medio interno a favor del gradiente de concentración. Lo curioso, sin embargo, es que el flujo osmótico de agua hacia el exterior no es tan intenso como cabría esperar de un gradiente tan fuerte (de unos 600 mOsm l-1). En principio podría pensarse que en los peces marinos el flujo de agua hacia el exterior del organismo es mucho mayor que el que experimentan los peces de río en sentido opuesto (hacia el interior), dado que en estos el gradiente osmótico con el medio externo es muy inferior (de unos 300 mOsm l-1). Sin embargo, ambos flujos son de magnitudes no muy diferentes si bien, como es lógico, de sentido contrario. La razón de que el flujo de salida de agua de los teleósteos marinos no sea demasiado intenso es que sus tegumentos han alcanzado un mayor grado de impermeabilización que los de los teleósteos de río y de esa forma compensan el efecto del mayor gradiente osmótico. Nos encontramos, por lo tanto, con que la impermeabilización del tegumento opera en los teleósteos marinos como barrera frente a la desecación, de la misma forma que operaba en los de agua dulce frente a la dilución.
El otro riesgo que han de conjurar los reguladores hiposmóticos es el de la entrada de sales en su organismo por difusión, dado que Na+ y Cl–, principalmente, y demás iones inorgánicos propios del agua de mar se encuentran mucho más concentrados en el medio externo que en el interno. El Na+, no obstante, tiene escasa tendencia a entrar o puede no tenerla, puesto que la cara interna del epitelio branquial –principal enclave por el que podría producirse su difusión- está cargada positivamente, por lo que el gradiente electroquímico no favorece la entrada. Ocurre lo contrario con el ión Cl–, dado que no solo el gradiente de concentración es favorable; también lo es el gradiente eléctrico. Existe, por lo tanto, una fuerte tendencia del Cl– a difundir hacia el interior.
El principal mecanismo que ponen en juego los teleósteos marinos para contrarrestar las pérdidas de agua consiste en beber. Al contrario que los de agua dulce, los marinos sí beben. En general, beben diariamente un volumen de agua que representa entre un 10% y un 20% de su masa corporal, aunque algunos no llegan al 1% diario y otros llegan al 50%. Ahora bien, de la misma forma que en los de agua dulce la producción de una orina copiosa generaba un problema de economía de iones inorgánicos (porque tienden a perderlos a través de la orina), en los marinos beber conlleva también serias complicaciones en lo relativo a la regulación de los flujos de sales, como veremos a continuación.
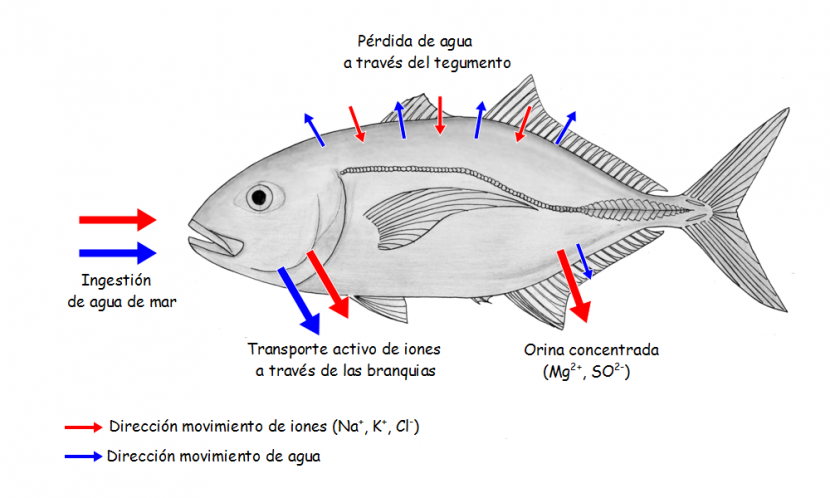
Al ingerir agua con alta concentración de sales inorgánicas, como la de mar, lo lógico es que esa agua no solo no sea absorbida, sino que, de hecho, el flujo osmótico se produzca en sentido contrario. Esto es, dado que la concentración salina y la osmótica total es muy superior en el contenido digestivo que en el plasma sanguíneo, cabe esperar que el agua fluya a favor de gradiente osmótico del medio interno a la luz intestinal. Y de hecho, eso es lo que ocurre durante buena parte del recorrido que sigue la ingesta a lo largo del sistema digestivo. Llega un momento, no obstante, en que el contenido intestinal aumenta y se diluye hasta que su concentración osmótica se iguala con la de la sangre. Bajo esas circunstancias se produce la absorción de agua. El agua se absorbe gracias a un proceso denominado transporte cuasi-isosmótico de fluidos que propicia su paso a la sangre, aun cuando los fluidos intestinales y el plasma son prácticamente isosmóticos. El movimiento de agua es posible gracias a la generación, mediante la absorción activa de iones monovalentes, de gradientes osmóticos locales en ciertas áreas del epitelio; parece ser que las acuaporinas del epitelio intestinal juegan un papel importante en ese proceso. De esa forma se puede llegar a absorber hasta un 85% (un 50% mínimo) del agua ingerida, aunque a cambio, casi todo el NaCl presente en ese agua es también absorbido, con lo que el problema que representa la regulación de las concentraciones iónicas se agrava. Los iones divalentes (SO4-2 y Mg+2, principalmente) se absorben en mucha menor medida que los monovalentes y son, en gran parte, eliminados con las heces.
Como hemos señalado antes y se ha podido comprobar, la regulación del contenido hídrico dificulta la regulación del contenido en sales de la sangre, puesto que para mantener el volumen de agua es necesario incorporar iones monovalentes. Es preciso, por lo tanto, eliminar esas sales. Los pocos iones divalentes absorbidos se eliminan en la orina que, en los teleósteos marinos, es isosmótica con el plasma y muy escasa (entre un 0,5 y un 3,5% del peso corporal por día). Hay dos poderosas razones para producir poca orina: (1) se pierde así poca agua por esa vía, y (2) dado que el agua que entra en estos peces está mucho más concentrada que el medio interno, si produjesen mucha orina, se retendría una cantidad excesiva de solutos en el plasma, solutos que habría que eliminar de otra forma.
Los iones monovalentes Na+ y Cl–, que han entrado del exterior a través del tegumento y desde el fluido intestinal, han de ser expulsados. Ya hemos visto que eso no ocurre a través de la orina. Son las branquias las encargadas de cumplir esa función. En concreto, son las células ricas en mitocondrias, también llamadas células de cloruro, las responsables de transportar el exceso de iones monovalentes al exterior. En aproximadamente la mitad de especies de teleósteos marinos, el transporte es exclusivamente de Cl–, porque el Na+ difunde al exterior a favor del gradiente eléctrico que genera el movimiento del anión. En otras especies ambos iones son transportados activamente. En los teleósteos marinos la eliminación de NaCl se produce, por lo tanto, de forma extrarrenal y es un proceso de gran relevancia cuantitativa, hasta el punto de que se puede afirmar que las branquias son el principal órgano osmorregulador de estos peces.
Los procesos de transporte activo de sales a que se ha hecho mención aquí son energéticamente costosos. La osmorregulación representa en los teleósteos marinos entre un 8% y un 17% del gasto metabólico basal. Eso es mucho. En los de agua dulce ese gasto es aproximadamente la mitad porque, como se dijo al comienzo, el gradiente osmótico es muy superior en los peces marinos, el doble aproximadamente que en los de agua dulce.
Sobre el autor: Juan Ignacio Pérez (@Uhandrea) es catedrático de Fisiología y coordinador de la Cátedra de Cultura Científica de la UPV/EHU
Los intercambios de agua y sales en los tetrápodos acuáticos – Cuaderno de Cultura Científica
[…] que tiene esta rana de resolver sus problemas osmóticos es mixta. Los renacuajos se comportan como los peces osteíctios marinos, regulando la concentración osmótica y salina de su medio interno y manteniéndolas en […]
Economía del agua en animales terrestres – Cuaderno de Cultura Científica
[…] vimos al estudiar los reguladores hiperosmóticos y los hiposmóticos, incluidos los tetrápodos, la producción de orina es una variable clave en la regulación […]
La función renal en vertebrados – Cuaderno de Cultura Científica
[…] atribuye al hecho de no necesitar producir grandes volúmenes de orina diluida. Los peces marinos son reguladores hiposmóticos y el riesgo principal que afrontan es el de la desecación. Además, y por la misma razón, estos […]