Formas moleculares de excreción de restos nitrogenados

Como apuntamos en la primera anotación de esta serie, la mayor parte de los animales eliminan sus restos nitrogenados a través de los sistemas de excreción y osmorregulación. En las anotaciones anteriores nos hemos ocupado de los mecanismos mediante los que esos sistemas producen la orina y la forma en que los tubos excretores mantienen los equilibrios hídrico y salino. Ha llegado el momento de referirnos de manera específica a las moléculas que se utilzan para eliminar los restos nitrogenados.
El metabolismo animal da lugar a la formación de productos tales como H2O y CO2. El agua metabólica pasa a formar parte de los elementos que intervienen en el balance hídrico, por lo que no ha de ser eliminada. El CO2, sin embargo, sí ha de ser expulsado, por las razones que expusimos en su día. Y el metabolismo de proteínas y ácidos nucleicos da lugar a la producción de restos nitrogenados.
Amonio
![]()
El producto terminal mayoritario del metabolismo de las sustancias nitrogenadas es el amoniaco (NH3), aunque en condiciones fisiológicas suele encontrarse en forma de ión amonio (NH4+). De hecho, lo más probable es que esta sea la molécula de excreción primitiva de los animales. Pero el amonio es una molécula muy tóxica. Entre otros efectos negativos, interfiere con el funcionamiento de la ATPasa de Na+/K+, lo que provoca una grave alteración de la integridad funcional de las membranas celulares. Esa es la razón por la que los efectos tóxicos del amonio se manifiestan, sobre todo, en el funcionamiento de las neuronas, que es muy dependiente de los fenómenos transmembrana. Concentraciones de amonio tan bajas como 2 mM en peces y 0,5 mM en mamíferos perjudican seriamente la función neuronal. Por esa razón el amonio ha de ser o bien eliminado o mantenerse muy diluido. Los animales acuáticos (en este caso ha de precisarse: animales que respiran en agua) no suelen tener mayores problemas. De hecho, no es raro que eliminen el amonio directamente a través de la pared corporal o de los epitelios branquiales. Así lo hacen la mayor parte de invertebrados marinos, peces teleósteos (tanto dulceacuícolas como marinos) y anfibios (larvas y adultos de vida acuática). A todos estos animales se les denomina amoniotélicos y a la forma de excreción, amoniotelismo.
Hay muy pocos animales terrestres que sean amoniotélicos, ya que para eliminar amonio hace falta mucha agua. De hecho, los pocos que eliminan amonio, o bien son animales de hábitats muy húmedos o lo hacen directamente en forma de NH3 gaseoso, como algunos isópodos terrestres. Algunos caracoles también, aunque no son amoniotélicos, eliminan parte de sus restos nitrogenados en forma gaseosa. Al parecer, esa forma de excreción está relacionada con la deposición de carbonato cálcico en el exoesqueleto.
Urea
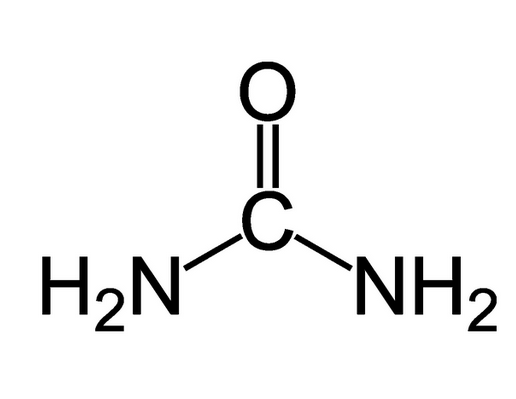
Otra molécula de excreción muy frecuente en el dominio animal es la urea. Es muy soluble y atraviesa con facilidad las membranas biológicas, aunque también tiene cierta toxicidad. De hecho, las concentraciones sanguíneas humanas se mantienen entre 3 y 7 mM, aunque pueden llegar a valores muy superiores. Por lo tanto, la excreción de restos nitrogenados en forma de urea necesita mucha menos agua que la excreción de amonio. En contrapartida, así como el amonio no requiere de un esfuerzo metabólico especial porque es el producto terminal que resulta de una secuencia de transaminaciones y desaminaciones de aminoácidos y sustancias similares, la urea ha de ser sintetizada y su síntesis requiere gasto de ATP (4 o 5 moléculas de ATP por molécula de urea). Este gasto es, pues, el coste que pagan los organismos que excretan sus restos nitrogenados en forma de urea a cambio de un importante ahorro de agua.
Ese ahorro explica el hecho de que la mayor parte de los animales que recurren a la urea como molécula mayoritaria de excreción sean vertebrados terrestres. Son ureotélicos los anfibios de vida adulta terrestre, algunas tortugas y todos los mamíferos. Sin embargo, apenas hay invertebrados que recurran al ureotelismo. Lo más probable es que este rasgo surgiera en los vertebrados que colonizaron el medio terrestre. Eso es lo que cabe deducir del hecho de que los peces de agua dulce –el grupo del que surgen los tetrápodos- sean mayoritariamente amoniotélicos. Del mismo modo, los anfibios terrestres transitan del amoniotelismo, característico de su fase larvaria acuática, al ureotelismo, propio de la vida adulta terrestre. La transición ocurre en paralelo a la metamorfosis y se produce al activarse –por efecto hormonal- la síntesis de las cinco encimas que catalizan las reacciones del ciclo de la urea. Los peces pulmonados y otras especies de peces también empiezan a sintetizar urea y dejan de producir amonio cuando se ven expuestos a estrés hídrico con carácter transitorio.
Como vimos aquí, además de ser usada como molécula de excreción, los condrictios y celacantos acumulan urea en su medio interno con fines osmóticos. Este dato indica que probablemente todos los vertebrados disponen de los genes necesarios para contar con un ciclo de la urea funcional; es la hipótesis que mejor explica el hecho de que la capacidad para sintetizar la molécula se encuentre tan extendida en este grupo y ocurra bajo circunstancias tan diversas.
Ácido úrico
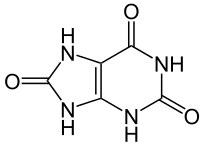
El ácido úrico y algunas formas químicas derivadas constituyen el tercer gran producto o grupo de productos de excreción de restos nitrogenados. Se suelen incluir en el grupo al ácido úrico, el ácido úrico dihidrato y los uratos, todas ellas purinas. Tienen una característica que las hace muy interesantes como moléculas de excreción: su baja solubilidad. Un litro de agua a 37ºC puede contener 0,4 milimoles de ácido úrico (lo que viene a ser 65 mg); por encima de esa concentración precipita. Por esa razón, en muchas ocasiones esas sustancias se eliminan en forma sólida o semisólida, de manera que se produce un gran ahorro de agua.
La baja solubilidad del ácido úrico es muy útil cuando un animal experimenta fuertes restricciones hídricas. Bajo esas circunstancias, en vez de aumentar su concentración, que es lo que ocurriría con la urea, una vez alcanzada la saturación, las cantidades adicionales precipitarían, acumulándose de forma sólida y, por lo tanto, evitando ejercer efectos tóxicos. Además del ácido úrico y los uratos, otras purinas, como la guanina o derivados del úrico, como la alantoína o el ácido alantoico, también son utilizados como molécula de excreción.
No hay acuerdo entre los especialistas acerca de los costes que conlleva la síntesis de ácido úrico. Algunos sostienen que son similares a los de la urea, aunque según otros pueden llegar a triplicar aquellos. No obstante, aunque la diferencia no llegue a ser tan importante, lo más probable es que, efectivamente, los costes sean mayores. Las ventajas que se derivan de su baja solubilidad, sin embargo, parecen compensar esa diferencia, por el gran ahorro de agua que conlleva su excreción. Eso explica que la mayor parte los animales terrestres sean uricotélicos. Lo son las aves, los lagartos, las serpientes, las tortugas que ocupan medios muy secos (ácido úrico), los arácnidos (guanina), la mayoría de insectos terrestres (ácido úrico, alantoína o ácido alantoico) y, en general, la mayoría de invertebrados terrestres.
Las reacciones metabólicas que posibilitan el uricotelismo son variantes de la vía de síntesis de las purinas, lo que explica el hecho de que hayan aparecido en tantos grupos animales. No está clara cuál es la razón por la que los mamíferos no son uricotélicos, aunque podría ser por la gran capacidad que han desarrollado para concentrar la orina en la urea gracias a su excepcional nefrona. Ello les permite un importante ahorro de agua a pesar de todo.
Para terminar, conviene reparar en el hecho de que todos los animales uricotélicos tienen desarrollo cleidoico, por lo que la posibilidad de almacenar úrico de forma sólida en el interior del huevo ha podido ser un factor determinante para la aparición del uricotelismo.
Sobre el autor: Juan Ignacio Pérez (@Uhandrea) es catedrático de Fisiología y coordinador de la Cátedra de Cultura Científica de la UPV/EHU
