La ventilación de los pulmones de mamíferos

La presión que ejerce un gas a una temperatura constante varía en función inversa del volumen que ocupa dicho gas. En otras palabras, si un número determinado de moléculas del gas pasa a ocupar mayor volumen del que tenía, la presión que ejerce disminuye. Y al contrario. En esa simplísima norma se basa el mecanismo que permite que fluya aire de la atmósfera a los pulmones y de estos al exterior. Me refiero a pulmones de mamíferos, aunque en realidad sirve para cualquier otro pulmón aunque las cosas ocurran de forma algo diferente.
En los pulmones de mamíferos el movimiento mareal (de entrada y salida) del medio respiratorio está provocado por cambios de volumen de la caja torácica. Ese volumen cambia debido a la contracción alterna de los músculos intercostales internos y externos, así como del diafragma y de los músculos abdominales. El aumento del volumen se produce como consecuencia de la contracción de los músculos intercostales externos y del diafragma; el esternón se eleva al separarse unas costillas de otras y, como consecuencia, se aleja de la columna vertebral. Al elevarse el volumen de la caja también lo hace el del interior pulmonar, por lo que la presión interna disminuye y, dado que la presión exterior no varía (760 mmHg a nivel del mar), el gradiente de presiones produce el flujo hacia el interior. La espiración se produce al relajarse el diafragma y contraerse los músculos intercostales internos y los abdominales. Se reduce así el volumen de la caja, con lo que aumenta la presión intra-alveolar y produce el flujo de aire hacia el exterior.
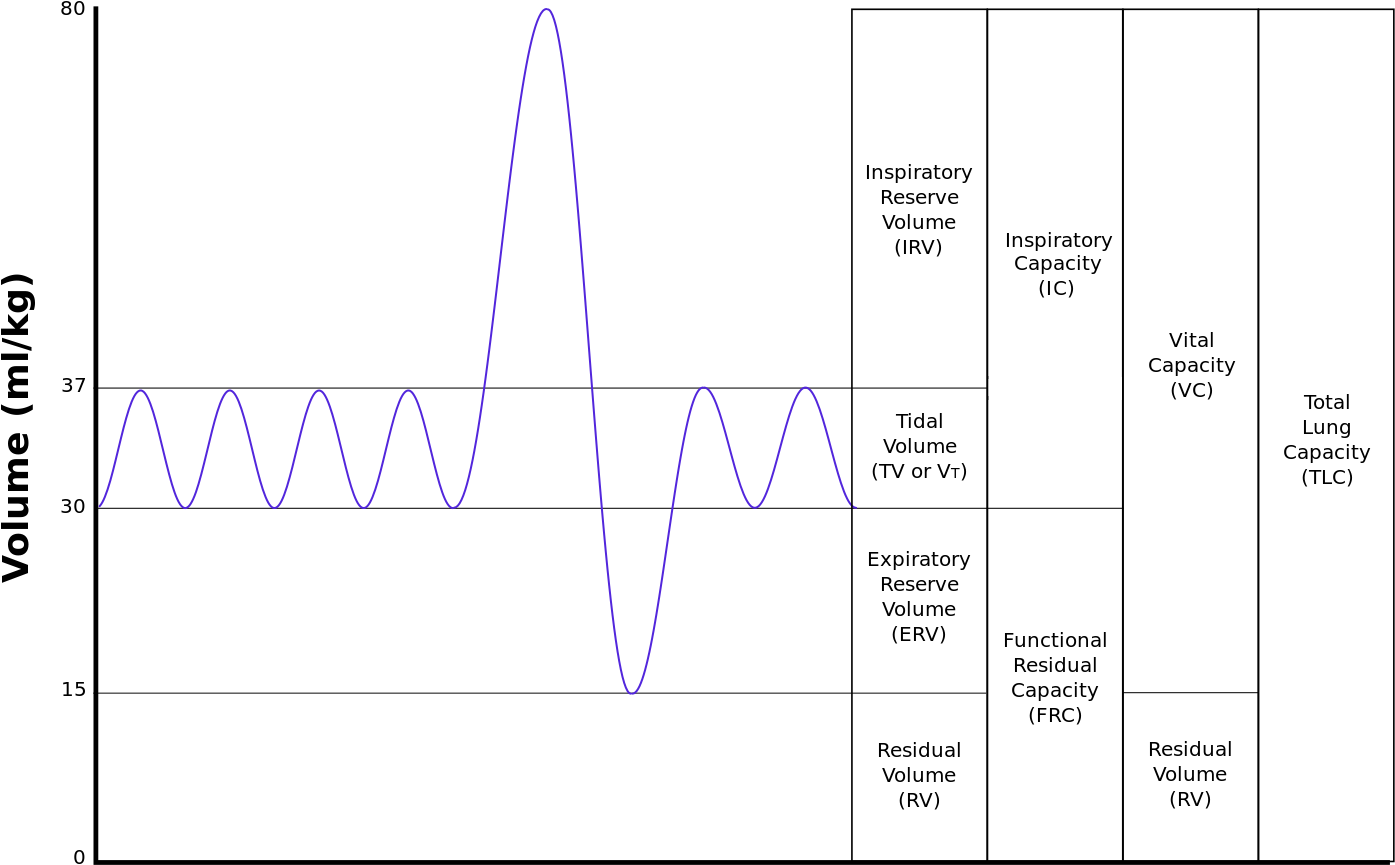
Los movimientos respiratorios en los pulmones de mamíferos definen una serie de volúmenes y capacidades que sirven para caracterizar el funcionamiento del órgano. La técnica que se utiliza para determinar esas magnitudes es la espirometría, realizada mediante un instrumento que mide el volumen de aire que es desplazado tras la ejecución de diferentes movimientos respiratorios. Se ofrecen a continuación esos indicadores junto con el valor de la media para un caballo y un hombre (las mujeres presentan valores algo más bajos):
- Volumen mareal (VM): es el volumen que entra o sale de los pulmones durante una única respiración. [6000 ml; 500 ml]
- Volumen inspiratorio de reserva (VIR): es el volumen extra de aire máximo que puede ser inspirado por encima del volumen mareal en condiciones de reposo. [12000 ml; 3000 ml]
- Capacidad inspiratoria (CI): es el máximo volumen de aire que puede ser inspirado al final de una espiración normal. CI = VM + VIR [18000 ml; 3500 ml]
- Volumen espiratorio de reserva (VER): es el volumen extra de aire que puede ser activamente espirado mediante la contracción máxima de los músculos espiratorios, en añadidura al que se espira normalmente de forma pasiva al final de una espiración normal típica. [12000 ml; 1000 ml].
- Volumen residual (VR): el volumen mínimo de aire que permanece en los pulmones tras una espiración máxima. [12000 ml; 1200 ml] Como es lógico, este volumen no puede establecerse por métodos espirométricos, puesto que ese aire no puede ser expulsado; se mide mediante técnicas de dilución con algún marcador gaseoso inocuo, como el Helio.
- Capacidad residual funcional (CRF): es el volumen de aire que queda en los pulmones al final de una respiración pasiva normal. CRF = VER + VR [24000 ml; 2200 ml]
- Capacidad vital (CV): es el máximo volumen de aire que puede desplazarse durante una única respiración tras una inspiración máxima. Se inspira y, a continuación, se espira todo lo que se puede. CV = VIR + VM + VER [30000 ml; 4500 ml]
- Capacidad pulmonar total (CPT): es el máximo volumen de aire que pueden albergar los pulmones. CPT = CV + VR [42000 ml; 5700 ml]
En resumen, en una respiración normal, tomamos y expulsamos 0,5 l de aire. En esas condiciones quedan en el interior de los pulmones 2,2 l. No obstante, estos pueden llegar a albergar 5,7 l; y dado que el máximo que pueden intercambiar en un único ciclo respiratorio es de 4,5 l, siempre quedarán dentro de los pulmones 1,2 l sin intercambiar. Prácticamente nunca se desplaza todo el volumen que podría intercambiarse como máximo en un único ciclo, por lo que, dependiendo de las necesidades, inspiramos entre 0,5 l y, en caso de gran necesidad, otros 4 l más.
De lo anterior se deriva que en los pulmones siempre hay un cierto volumen de aire con oxígeno (el volumen residual), por lo que incluso durante la espiración y antes de que vuelva a tomarse aire, la transferencia de oxígeno a la sangre no cesa por lo que solo se producen pequeñas variaciones en la concentración sanguínea de este gas al salir de los pulmones.
Por último, merece la pena reparar en la existencia de un volumen de aire que no es fisiológicamente útil. Me refiero al del espacio muerto, un volumen que no participa en el intercambio de gases respiratorios. Ese espacio da cuenta de unos 150 ml. Es el aire que permanece en los bronquios en zonas donde no se produce intercambio de gases o que accede a alveolos mal perfundidos. Para valorar las implicaciones de la existencia de ese espacio muerto, hagamos unas cuentas sencillas.

En condiciones normales respiramos con una frecuencia de 12 min-1 (doce movimientos inspiratorios y otros tantos espiratorios por minuto) y en cada uno intercambiamos 500 ml de aire. Por lo tanto, la ventilación pulmonar es de 6 l min-1. Sin embargo, no todos los 500 ml participan en el intercambio, ya que 150 ml corresponden al espacio muerto. Así pues, la ventilación alveolar –la efectiva a efectos respiratorios- es de 4,2 l min-1. Por ello, en una situación de necesidad es más rentable elevar el volumen mareal (volumen de intercambio) que la frecuencia respiratoria. Al subir el volumen mareal la importancia relativa del espacio muerto disminuye, pero al aumentar la frecuencia respiratoria no. Valoraré ahora tres supuestos al objeto de ilustrar esa afirmación.
El primer supuesto corresponde al ejemplo visto antes: una persona que respira con normalidad, con un volumen mareal de 500 ml y una frecuencia respiratoria de 12 min-1; su ventilación pulmonar es, como hemos visto, de 6 l min-1 y su ventilación alveolar de 4,2 l min-1. Si esa misma persona decide respirar muy rápidamente (40 min-1) y, por ello, hacerlo de forma superficial, podríamos encontrarnos con que su volumen mareal sea de 150 ml; en ese caso, su ventilación pulmonar sería igual que la anterior (6 l min-1), pero su ventilación alveolar sería nula (0 l min-1), pues el volumen mareal es igual al espacio muerto. En otras palabras, solo intercambiaría con el exterior el aire que se encuentra en las vías respiratorias donde no se produce intercambio de gases. Por el contrario, si optase por reducir su frecuencia respiratoria (a 5 min-1, por ejemplo), pero respirando profundamente (con un volumen mareal de 1,2 l, por ejemplo), la ventilación pulmonar seguiría siendo de 6 l min-1, pero la alveolar sería de 5,25 l min-1. Queda claro, por lo tanto, que la modalidad de respiración (somera, normal o profunda) ejerce un claro efecto sobre la ventilación alveolar que es lo que, a fin de cuentas, importa. Las aves, como vimos en otra ocasión, no tienen estos problemas, porque el aire circula de forma continua en un solo sentido y todo el que pasa por los bronquios participa en el intercambio de gases respiratorios. De ahí su extraordinaria eficacia.
Sobre el autor: Juan Ignacio Pérez (@Uhandrea) es catedrático de Fisiología y coordinador de la Cátedra de Cultura Científica de la UPV/EHU
Hitos en la red #189 | Enlace Recomendado | Naukas
[…] Sistemas respiratorios: la ventilación de los pulmones de mamíferos […]
gabriela
Profesor, quiero darle las gracias por esta información, que me ha servido muchísimo para entender mi problema de salud y la espirometría.