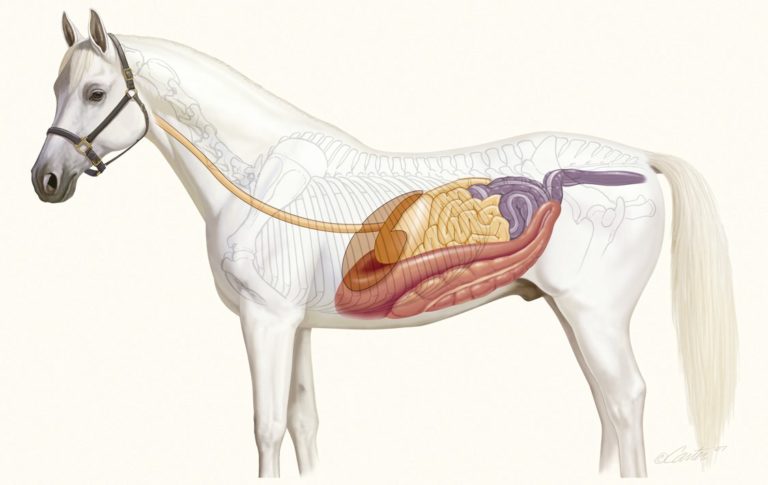
El intestino grueso
El intestino grueso de los vertebrados sigue al intestino delgado. Consta de tres partes principales: ciego, colon y recto (o cloaca). El ciego es una especie de bolsa que, como su propio nombre indica es ciego, o sea, es un fondo de saco. Recibe el quimo del íleo, la parte final del intestino delgado, aunque ciego e íleo se encuentran separados por la válvula ileocecal, que evita posibles retrocesos de los jugos intestinales. El íleo desemboca en un lateral del ciego y algo por debajo de ese punto se encuentra el apéndice, un tejido linfoide que no toma parte en la digestión, pero que contiene linfocitos.
El colon es la parte más extensa del intestino grueso y su estructura varía mucho de unas especies a otras en función de la dieta. Y dentro de los mamíferos también se da una gran variabilidad. En la especie humana se distinguen cuatro partes: el colon ascendente, el transverso, el descendente y el sigmoide.
En anfibios, reptiles (incluidas las aves) y algunos mamíferos, el intestino grueso finaliza en una cloaca en la que confluye con el sistema excretor y con el aparato reproductor. En los peces, sin embargo, los conductos de los tractos excretor y reproductivo salen al exterior de manera independiente del sistema digestivo. En reptiles (aves incluidas) unos pliegues o crestas delimitan la cloaca en tres áreas diferenciadas: coprodeo (anterior), a donde llegan las heces desde el intestino; urodeo (medio), que recibe la orina de los riñones a través de los uréteres y los materiales del oviducto; y el proctodeo (posterior), donde se almacenan las excretas. El proctodeo se abre al exterior a través de un ano muscular.
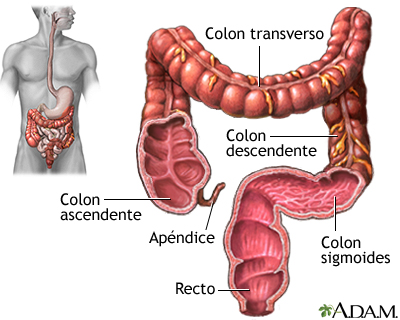
Un colon humano recibe diariamente del orden de medio litro de quimo. Cuando llega procedente del intestino delgado, los nutrientes han sido digeridos y absorbidos prácticamente en su totalidad, por lo que contiene sobre todo residuos alimenticios indigeribles (como celulosa, por ejemplo), algunos componentes biliares que no se han absorbido y, sobre todo, agua y sales. Por esa razón, en el colon prosigue la absorción de agua, que tiene lugar gracias a la recuperación activa de sodio, que va acompañada por la correspondiente reabsorción (a favor de gradiente eléctrico) de cloro; el agua pasa a favor del gradiente osmótico generado por la reabsorción de ClNa.
El colon contiene una solución mucosa alcalina (HCO3– y PO43-) que tiene efecto tamponador. Esa solución protege la mucosa del intestino grueso del daño mecánico y químico que podría sufrir si careciese de ella. En caballos y cerdos basta con el HCO3– de origen pancreático para mantener tamponado el colon. La saliva de los rumiantes contiene cantidades importantes de PO43-, pero en los demás mamíferos el fosfato intestinal es de origen alimenticio. Los tampones neutralizan los ácidos producidos por la fermentación bacteriana (a la que nos referiremos en una anotación próxima), y el moco proporciona la lubrificación que facilita el paso de los contenidos intestinales y la expulsión final de las heces.
Las dimensiones y estructura de los sistemas digestivos y, en especial, del intestino grueso depende mucho de la dieta propia de cada especie. Los peces carnívoros, por ejemplo, tienen tractos digestivos cuya longitud es solo algo mayor que la longitud corporal; los herbívoros, sin embargo, pueden llegar a ser veinte veces más largos.
Los vertebrados carnívoros tienen tubos digestivos muy simples, y el colon suele ser corto y sin áreas diferenciadas. Apenas se diferencian el intestino delgado y el intestino grueso; no suelen tener ciego, y si lo tienen, es marginal. En estas especies la función principal (casi única) del intestino grueso es la absorción de sales y agua.
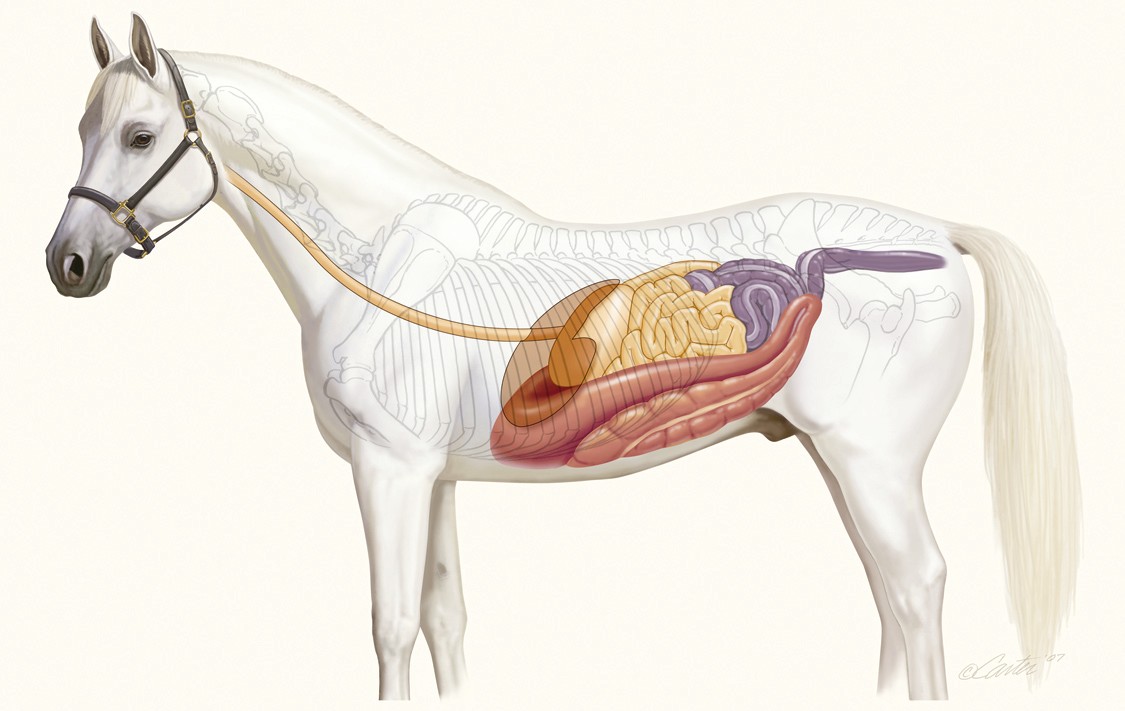
En los omnívoros y, sobre todo, en los herbívoros, la estructura del intestino grueso tiende a ser más compleja; conejos y caballos son buenos ejemplos. El ciego de los herbívoros suele ser grande, en algunos casos mayor que el colon, incluso. Además, tanto el ciego como el colon suelen estar provistos de saculaciones (sacos laterales expandibles). No obstante, hay excepciones: ni canguros ni ovejas se caracterizan por tener colon y ciego de grandes dimensiones, ni por tener saculaciones.
Las saculaciones se forman cuando la capa exterior de músculo liso longitudinal no envuelve el intestino grueso por completo. En las especies cuyos intestinos gruesos presentan esas estructuras, hay bandas de musculatura lisa longitudinal denominadas taeniae coli (en singular taenia coli), que discurren todo a lo largo del ciego y el colon. La mayoría de los animales que las tienen cuentan con tres, aunque el caballo, por ejemplo, tiene cuatro. Son más cortas que lo que las capas muscular (circular) y mucosa subyacentes serían de no estar constreñidas en parte por las taeniae coli. La presencia de estas bandas musculares provoca, por ello, que se formen leves constricciones que, a su vez, delimitan los embolsamientos o saculaciones a los que denominamos haustras (haustra coli). Los haustras no son estructuras estáticas, sino que cambian de posición por efecto de las contracciones de la capa circular de musculatura lisa.
Las contracciones haustrales juegan un papel primordial en la motilidad del intestino grueso. Se inician debido a la actividad rítmica autónoma de las células musculares lisas del colon, y son similares a las contracciones que provocan la segmentación en el intestino delgado, solo que se producen con una frecuencia muy inferior. Se producen entre 9 y 12 contracciones de segmentación por minuto y, sin embargo, entre dos contracciones haustrales pasan del orden de 30 minutos. Como consecuencia de esa actividad contráctil, la localización de los sacos haustrales cambia, porque las zonas que se contraen y las que se relajan se van alternando. Esta motilidad no provoca desplazamiento unidireccional del contenido intestinal; su función es ir exponiéndolo gradualmente a la acción absortiva del epitelio. Y al no provocar su avance, permanece largo tiempo en el interior del intestino grueso, de manera que se prolonga el tiempo en que se produce la acción de las bacterias intestinales sobre los materiales de origen vegetal que no habían sido digeridos y absorbidos en el intestino delgado.
Además de la provocada por las contracciones haustrales, en el intestino grueso se producen otros dos tipos de motilidad. Por un lado están las contracciones peristálticas; son provocados por células marcapasos ubicadas en la zona media del colon y generan ondas lentas en las dos direcciones. Su efecto neto consiste en impulsar el contenido digestivo hacia el recto. La motilidad intestinal de las aves se basa, sobre todo, en contracciones peristálticas. Otras células marcapasos ubicadas en la región proximal del colon generan contracciones antiperistálticas, cuya principal función consiste en llenar el ciego. Son contracciones importantes en la mayoría de los herbívoros y en los cerdos, pues su acción de mezcla facilita la digestión bacteriana de la celulosa y la absorción de los ácidos grasos volátiles, producto de la fermentación.
Tras cada comida suele producirse un fuerte aumento de la motilidad del colon. Grandes segmentos se contraen de forma simultánea, desplazando el contenido intestinal entre un tercio y tres cuartas partes de su longitud en unos pocos segundos. Estos movimientos “en masa” (así se llaman), conducen el contenido del colon hacia la porción distal del intestino grueso, donde queda almacenado hasta su expulsión.
Cuando el alimento entra en el estómago, se producen los reflejos gastroileal y gastrocólico, en los que interviene la gastrina y nervios autónomos extrínsecos. El reflejo gastroileal provoca el vaciado del intestino delgado y el gastrocólico el desplazamiento del contenido del colon al recto. De esa forma se va haciendo hueco, primero en el intestino delgado y después en el ciego y el colon, a nuevos contenidos.
Cuando los movimientos en masa del colon llevan el material fecal al recto, la distensión a que da lugar estimula los receptores de estiramiento de la pared rectal, iniciando así el reflejo de defecación. Ese reflejo provoca la relajación del esfínter anal interno (musculatura lisa) y contracciones fuertes del colon sigmoide y del recto. El esfínter externo consiste en musculatura esquelética y, por lo tanto, controlada voluntariamente; por lo tanto, si se relaja ese esfínter también, se produce la defecación. No obstante, si el esfínter exterior permanece contraído de manera prolongada, las contracciones rectales cesan hasta que se produce la llegada de material fecal adicional por efecto de posteriores movimientos en masa, y se reinicia el reflejo de defecación.
Las heces está formadas por celulosa que no se ha digerido y otras sustancias que no se han podido absorber, como bilirrubina (o biliverdina), sales, agua y bacterias. Aunque en el colon tiene lugar una cierta absorción, su importancia cuantitativa es mínima comparada con la del intestino delgado. La superficie interna del intestino grueso es muy reducida por comparación con la del delgado, dada la ausencia en aquel de las estructuras que multiplican la superficie absortiva propias de este. Además, el colon de muchos vertebrados carece de mecanismos de transporte específicos para las sustancias que se absorben en el intestino delgado. La excepción es el colon de las aves, en el que se absorbe glucosa y aminoácidos mediante transporte secundario activo. De esa forma, los aminoácidos y la glucosa que no han sido recuperados de la orina por el riñón pueden ser reabsorbidos en el colon o ser utilizados por los microorganismos en los ciegos intestinales.
Sobre el autor: Juan Ignacio Pérez (@Uhandrea) es catedrático de Fisiología y coordinador de la Cátedra de Cultura Científica de la UPV/EHU