Digestión simbiótica: los rumiantes

La fermentación simbiótica está muy extendida en el reino animal y casi todos los vertebrados, en especial los herbívoros y omnívoros, tienen alguna forma de simbiosis digestiva de ese tipo. La que ha alcanzado en estos un mayor grado de sofisticación es la de los rumiantes. Pero no es la única. De hecho, y aunque los casos en los que aparecen estructuras especializadas como el rumen han sido los más estudiados, durante los últimos años la microbiota intestinal de otros mamíferos y, muy en especial, la humana han recibido una gran atención. Aquí, no obstante, nos limitaremos a repasar de forma somera aquellos casos en los que la fermentación simbiótica se produce en estructuras especializadas o implica comportamientos alimenticios específicos.
Cuando utilizamos el término fermentación en este contexto nos referimos a un conjunto de reacciones enzimáticas que tienen lugar en entornos carentes de oxígeno y que dan lugar a la ruptura de compuestos orgánicos que rinden moléculas energéticas que pueden ser utilizadas en el metabolismo del animal hospedador.
Las estructuras especializadas que albergan microbios suelen ser cámaras en las que el flujo de materiales a su interior propicia un ambiente adecuado (húmedo o semilíquido, cálido y no excesivamente ácido) para el crecimiento microbiano. En algunas especies esas cámaras se encuentran en el sistema digestivo anterior, la parte comprendida por el esófago y el estómago; a esas especies se las denomina fermentadoras pregástricas. Los más conocidos son los mamíferos rumiantes. También hay fermentadores posgástricos y, en algunos invertebrados, otras modalidades de simbiosis digestiva.
De acuerdo con los modelos matemáticos que se utilizan para simular el funcionamiento de los sistemas digestivos, la ruptura de las moléculas de alimento mediante fermentación microbiana es más efectiva cuando el material recién ingerido se mezcla con alimento ingerido antes, ya colonizado por bacterias y parcialmente digerido. Por el contrario, la digestión enzimática del alimento –la realizada por las enzimas producidas por el propio animal- es más efectiva cuando las enzimas se añaden a un material que no se ha mezclado aún con otro sometido previamente a digestión enzimática. A esa diferencia entre las eficiencias de un tipo y otro de digestión parece obedecer el hecho de que las cámaras especializadas para la fermentación microbiana sean amplias y tengan forma de tina o barril, mientras que el intestino es tubular, como en el resto de mamíferos.
Los rumiantes reciben su nombre del acto de rumiar (o rumia), que consiste en masticar la comida durante periodos prolongados de tiempo. Puede tratarse de alimento recién ingerido o de material regurgitado para someterlo a un segundo tratamiento en la boca. Los rumiantes han desarrollado un estómago de gran tamaño y motilidad, que les permite albergar grandes poblaciones microbianas que se ocupan de digerir celulosa y otros carbohidratos complejos, rindiendo productos finales útiles para el hospedador. La rumia ha sido un hallazgo impresionante, gracias al cual los animales que la han desarrollado han alcanzado un gran éxito evolutivo.
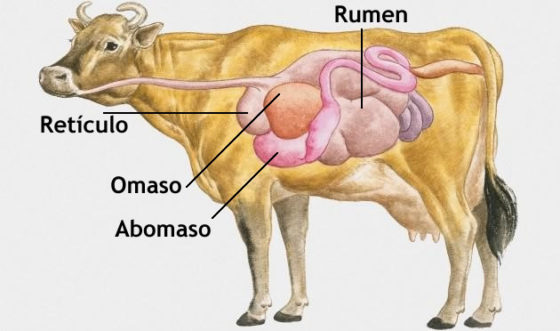
Los verdaderos rumiantes son los bovinos, ovinos, caprinos, cérvidos, jirafas y antílopes. Su estómago, que ocupa tres cuartas partes de la cavidad abdominal, está dividido en cuatro compartimentos: rumen (también llamado panza o herbario), que es el más espacioso; retículo (redecilla o bonete); omaso (libro o librillo); y abomaso (cuajar). Los tres primeros constituyen lo que se denomina estómago anterior o región pregástrica. Y el abomaso sería el verdadero estómago. Los pseudorrumiantes carecen de omaso; en este grupo se encuentran hipopótamos y camélidos. Los perezosos, canguros, monos colobos (mamíferos) y el hoazín (ave de las selvas americanas) también son fermentadores pregástricos, pero no son rumiantes.
Los tres compartimentos pregástricos conducen y almacenan alimento. El rumen y el retículo, además, también absorben nutrientes y moléculas pequeñas. El grueso de la fermentación simbiótica ocurre en esos dos compartimentos, y en ellos se mantienen las condiciones de temperatura, pH y motilidad idóneas para poder mantener las poblaciones microbianas que se ocupan de ella.
La cavidad del rumen está dividida en compartimentos internos –los sacos dorsal y ventral- mediante estructuras longitudinales denominadas pilares. Cuando se contraen, esas estructuras facilitan la mezcla de sus contenidos. Y también contribuyen a estabilizarlos limitando sus movimientos y evitando desplazamientos importantes de volúmenes relativamente grandes. Unas proyecciones de aspecto digital denominadas papilas recubren el interior del rumen, y le proporcionan una mayor área superficial, lo que facilita la absorción de nutrientes. Aunque el rumen y el retículo se encuentran parcialmente separados por el pliegue ruminorreticular, ambas cavidades no se diferencian demasiado y hay bastante intercambio de productos digestivos entre ellas.
El esófago termina en el cardia, donde retículo y rumen se conectan. La superficie interna del retículo se encuentra recubierta por crestas o rugosidades, de las que se cree realizan una cierta selección de las partículas que pasan cerca del orificio que une el retículo al omaso. Este conecta el retículo con el estómago glandular (abomaso) y en su pared interior hay unas estructuras similares a hojas por donde se produce absorción de agua y nutrientes, y que impiden el paso al abomaso de partículas de tamaño excesivamente grande. El orificio omasoabomasal conecta ambas cámaras; carece de esfínter que evite el retroceso de los contenidos digestivos. El abomaso es muy similar al estómago de los no rumiantes. Es donde ocurre la parte más importante de la digestión de proteínas y la lisis de las bacterias procedentes del rumen.
Los nervios vago (sistema parasimpático) y esplácnico (sistema simpático) –del sistema autónomo– inervan el estómago de los rumiantes. Las fibras motoras del nervio vago procedentes de los centros gástricos de la médula oblonga del tronco encefálico pueden provocar una mayor frecuencia de contracciones del retículo-rumen. Eso ocurre a la vista de alimento, al masticar o al rumiarlo. Los aumentos de la distensión del retículo-rumen también provocan un aumento de la velocidad a la que se contrae. Por el contrario, el efecto de la división simpática (nervio esplácnico) es inhibidor. La distensión del abomaso disminuye la tasa de contracción del retículo-rumen.
La rumia consiste en la regurgitación, remasticación y reingestión del alimento. El alimento nuevo se mezcla con saliva y es masticado en la cavidad bucal, de donde pasa, a través del esófago, a la cavidad retículo-rumen. Allí es degradado, mediante procesos de fermentación, por los microorganismos que contiene. La fracción más líquida y en la que se encuentran las partículas de menor tamaño sigue su curso a través del resto de estructuras del sistema digestivo. Pero el bolo, formado por los fragmentos más sólidos, es regurgitado a la boca donde vuelve a ser masticado dando lugar a un material más finamente triturado y, por lo tanto, con una mayor área superficial, lo que facilitará la digestión a cargo de los microbios cuando vuelve a ser ingerido y conducido de nuevo al retículo-rumen.
Bacterias, principalmente (se han identificado más de 200 especies), pero también algunos protozoos, arqueas y hongos degradan la celulosa y hemicelulosa, mediante celulasas, produciendo ácidos grasos de cadena corta. Las proteínas y los carbohidratos no estructurales (pectina, almidones y azúcares) también sufren la fermentación. La mayor parte de los carbohidratos simples que resultan de estas fermentaciones son utilizados por los microorganismos, aunque algunos pueden escapar del rumen y ser absorbidos después por el animal. En este proceso la saliva es muy importante, porque proporciona el medio líquido adecuado para que se desarrollen florecientes poblaciones microbianas, además actúa como tampón del pH del rumen, gracias a su contenido en bicarbonatos y fosfatos.
La degradación de los carbohidratos complejos por los microorganismos da lugar, a través de la glucolisis, a fosfoenol piruvato (PEP), del que se produce metano, CO2, acetato y algo de butirato. O también puede ser metabolizado a piruvato que, en última instancia, conduce a la formación de propionato y butirato. Bajo condiciones normales, el rumen contiene un 60-70% de acetato, un 14-20% de propionato y un 10-14% de ácido butírico. Estos ácidos de cadena corta son muy importantes para el animal. El propionato es el único que puede utilizar para sintetizar glucosa y glucógeno; bajo condiciones normales, el 70% de la glucosa de un rumiante proviene del ácido propiónico. Los otros ácidos grasos volátiles se incorporan al ciclo de Krebs como acetil CoA, y si hay excedentes, se acumulan en forma de grasas. Estos ácidos se absorben de forma pasiva a través de la pared ruminal.
Los microbios simbiontes hidrolizan las proteínas contenidas en el alimento y como consecuencia de esa hidrólisis se producen polipéptidos y aminoácidos que utilizan para su propio crecimiento. Casi todas las proteasas bacterianas se encuentran en su interior (el de las bacterias), pero algunas las liberan al exterior (a la luz del rumen) y actúan allí. En cualquier caso, los oligopéptidos de hasta 6 átomos de carbono son absorbidos por las bacterias y sometidos a ulteriores degradaciones hasta rendir aminoácidos o, incluso, amonio (por desaminación de aquellos). Las paredes del rumen pueden absorber con facilidad los aminoácidos y el amonio que no es utilizado por los propios microorganismos.
La mayor parte del amonio, así como algunos aminoácidos y oligopéptidos, son utilizados por los microorganismos para elaborar sus propias proteínas. Y una vez que llegan al abomaso, esas bacterias son atacadas y sus proteínas digeridas por los jugos estomacales. Los aminoácidos resultantes son absorbidos en el intestino delgado.
El poco amonio que no es reutilizado por las bacterias del retículo-rumen se absorbe y es convertido en urea1 en el hígado del rumiante, y parte de esa urea, junto con la que procede de la degradación de proteínas propia de los tejidos del animal, es transportada de nuevo al rumen, directamente a través de su pared o por las glándulas salivares. La alta actividad ureasa propia de la pared del rumen garantiza una rápida conversión de la misma en amonio para su utilización por la microbiota ruminal. Aquí radica gran parte de la importancia de la rumia y la fermentación pregástrica: además de facilitar el aprovechamiento de carbohidratos complejos, permite utilizar las proteínas de los alimentos con una eficiencia enorme, dado que solo una pequeña parte del nitrógeno ingerido se acaba evacuando en forma de urea.
Por último, conviene destacar que, como consecuencia de la fermentación, los microorganismos ruminales también producen todas las vitaminas B, incluido el complejo B12 si hay suficiente cobalto.
Nota:
1 A la urea, junto con las otras formas moleculares de excreción de nitrógeno nos referimos aquí.
Sobre el autor: Juan Ignacio Pérez (@Uhandrea) es catedrático de Fisiología y coordinador de la Cátedra de Cultura Científica de la UPV/EHU

RUIZ CHATEZ DANYERLY JOHANA
Hola, podrian regalarme la referencia bibliografica o la cita para esta pagina. Es para anexarla a mi trabajo.
Muchas gracias.
Nitroplasto, un nuevo orgánulo generado por endosimbiosis
[…] (bacterias y arqueas), deben su propia existencia y su evolución a la endosimbiosis. La simbiosis es la asociación de organismos con beneficio mutuo. Hablamos de endosimbiosis cuando uno de estos […]